Chapter 9 Fundamentals of Regenerative Medicine and Its Applications to Electrophysiology
The adult human heart has long been accepted as an end organ having no regenerative properties. In contrast, nonmammalian species such as zebrafish recover completely after ventricular apical resection thereby manifesting cardiac regeneration.1 Regenerative medicine builds on such observations, with the aim to replace or regenerate cells, tissues, and organs to restore or establish normal function.
Despite previous wisdom, recent evidence suggests that (1) adult human cardiomyocytes have mitotic potential, (2) cardiac progenitor cells can be isolated, (3) cardiomyocyte turnover occurs, and (4) human embryonic stem cells can differentiate into cardiomyocyte-like cells in culture.2–7 These discoveries have sparked excitement about the idea of repopulating the heart with healthy cardiomyocytes after a myocardial infarction, an idea that only recently was considered science fiction.4 Part of the effort in regenerative medicine has focused on cardiac arrhythmias. This effort has drawn on knowledge of the molecular and biophysical properties of the ion channels and signaling molecules that contribute to the initiation and propagation of the action potential. The effort has gained impetus from continued disappointment with the performance of antiarrhythmic drugs.
Gene Transfer by Viral Vectors
A successful gene therapy strategy must be safe, easy to deliver, and predictable in expression, efficacy, and duration of effect. Viral vectors have been widely used for gene transfer. Several factors need to be considered in choosing a viral vector: (1) the size of the gene it can incorporate; (2) the ease of genetic manipulation; (3) the ability to infect the target cell type; (4) replication deficiency; (5) lack of inflammatory and oncogenic potential; and (6) reliability of expression. Adenovirus and adeno-associated virus are favored for proof of concept studies because they are easily manipulated and have high expression levels. Durability of expression is the major limitation of adenovirus. Adeno-associated virus can mediate expression in the heart for months, if not longer, but the packaged gene size is limited to under five kilobases. Retroviruses such as lentivirus are incorporated into the genome and have the potential for long-term expression of the therapeutic gene.8–10
Stem Cell–Based Therapy
The multipotency of progenitor cells and pluripotency of embryonic stem cells are the fundamental properties that make them attractive for regenerative medicine, but they also raise safety questions. Undesirable differentiation and proliferation of stem cells may cause tumor formation. Also, stem cells migrate and home to specific biochemical signals. These homing properties can be exploited to target them to specific areas of disease.11 Alternatively, stem cells may migrate and lose their effect if they detect a chemoattractant located elsewhere. While autologous progenitor cells are ideal for limiting rejection, some stem cells (e.g., mesenchymal stem cells) appear to be immunoprivileged and have potential application in allogeneic therapy. Genetic engineering of stem cells can be accomplished by various techniques that use viruses, electroporation, or liposomes. Stem cells also represent a platform for “designer” therapeutics.
Treatment Strategies for Bradyarrhythmias: Biologic Pacemakers
Properties of an Ideal Biologic Pacemaker
Advances in microcircuitry and battery technology have miniaturized the modern electronic pacemaker such that implantation is now a routine procedure done outside a surgical operating suite. However, electronic pacemaker therapy has some shortcomings, such as the requirement for permanent hardware implantation, limited battery life, potential for malfunction, and a foreign body that may serve as a nidus for infections. The extraction of an infected pacemaker (especially an infected lead) is a complex undertaking that has a significant risk of mortality. The placement of pacemaker leads and the activation of the myocardium may impact unfavorably on cardiac contractility and electrophysiology. Furthermore, electronic pacemakers are not responsive to autonomic stimulation, especially that related to a physical activity or an emotional state. In the pediatric patient, electronic pacemaker hardware must be selected taking physical growth into consideration. Lastly, the function of the electronic pacemaker is prone to interference from common consumer electronic devices as well as medical equipment such as magnetic resonance imaging (MRI) equipment. These limitations have led to interest in the development of biologic pacemakers.12,13
Strategies to Create Biologic Pacemakers
Gene Therapy
Overexpression of β-Adrenergic Receptors
Glycoprotein (G-protein)–coupled β-adrenergic receptors regulate chronotropic and ionotropic responses to circulating catecholamines. The first successful gene transfer experiment resulting in biologic pacemaking used plasmids to overexpress the human β2-adrenergic receptor in the murine atrium.14 Edelberg et al then demonstrated that gene transfer was feasible by using catheter-based injection of plasmid into the procine right atrium. Overexpressing the human β2-adrenergic receptor was shown to increase heart rate by about 50% 2 days after plasmid injection.15 Further application of these studies was limited because the plasmid-based gene delivery system conferred only short-lived expression.
Inhibition of Diastolic Repolarization Current, IK1
Mutations within the pore regions of channels can dramatically affect channel conductance. Miake et al introduced an adenovirus packaged with the Kir2.1AAA mutant and green fluorescent protein (GFP) into the guinea pig left ventricle cavity.16 Transfected myocytes showed 80% suppression of IK1. The Kir2.1AAA-expressing myocytes exhibited two electrophysiological behaviors: (1) They lacked spontaneous activity with elicited prolonged action potentials, or (2) they expressed spontaneous activity remarkably similar to that of sinoatrial pacemaker cells. The electrocardiograms (ECGs) of transfected animals showed that half of them remained in sinus rhythm with QT prolongation, and the other half showed spontaneous ventricular rhythms that were at times faster than sinus.
Overexpression of If
Ion channels encoded by the HCN (hyperpolarization-activated, cyclic nucleotide–gated) gene family underlie the pacemaker current, If, which initiates depolarization during phase 4 of the sinoatrial action potential (Figure 9-1). Because If only activates on hyperpolarization, it does not have the potential to prolong the duration of the action potential and initiate proarrhythmia on this basis. Qu et al injected adenovirus carrying the mouse HCN2 gene (one of four HCN isoforms), into the canine left atrial appendage.17 A spontaneous cardiac rhythm originated from the left atrium in all four dogs studied during sinus arrest (induced by right vagal stimulation). Patch-clamping of isolated HCN2-expressing atrial myocytes showed If current magnitude 500 times greater than that in control atrial myocytes.
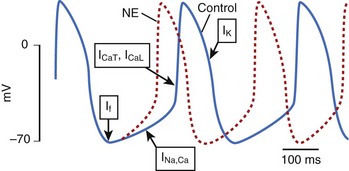
Plotnikov et al used catheter-based endocardial injection to deliver the adenovirus expressing HCN2 into the canine proximal left bundle branch, an ideal site for providing organized left ventricular activation when the distal conducting system is functional.18 Two days later, a left ventricular rhythm was observed during sinus arrest induced by vagal stimulation. Subsequently, stable pacemaker function was demonstrated following HCN2 injection into the left bundle branch of dogs with complete heart block.19 Expression with this adenoviral construct lasted 2 weeks.
Ion Channel Mutations
Structure-function studies of HCN2 channels have revealed that certain amino acids are critical to defining the channel’s operating characteristics. A point mutation (glutamic acid to alanine at position 324, E324A) in mHCN2 positively shifted the voltage dependence of activation and deactivation gating kinetics. The positive shift in voltage dependence of E324A channels generates a faster pacemaker rate and increased sensitivity to catecholamines than native HCN2. When adenovirus expressing E324A was injected into the canine left bundle branch, dogs receiving E324A were significantly more responsive to catecholamines.19 During epinephrine infusion, all E324A-injected dogs had their heart rate increase by at least 50%, whereas only a third of the HCN2-injected dogs and a fifth of the control dogs had a similar response. The E324A study also illustrates that gene therapy is not limited to using endogenous genes but that mutations can be tailored to function.
A chimeric approach to creating an HCN-based biologic pacemaker with faster basal rates was undertaken by Plotnikov et al.20 A channel with the N and C terminals of HCN2 and the transmembrane domains of HCN1 was created (HCN212) that would have HCN2’s superior catecholamine response with the favorable activation kinetics of HCN1. The HCN212 chimera had similar electrophysiological characteristics to HCN2 when expressed in isolated ventricular myocytes; however, the mean time constant of activation was faster in HCN212. An HCN212-based biologic pacemaker would likely result in a faster basal rate than an HCN2-based one, as more current would pass earlier during diastolic depolarization. Expression of HCN212 into the left bundle of dogs with complete heart block resulted in rapid ventricular tachycardia originating from the adenoviral injection site that was responsive to the If-blocking drug, ivabradine. Additional work to fine tune an HCN-based biologic pacemaker is needed. If If-associated arrhythmias occur with HCN-based pacemakers, If-blocking drugs maybe useful in the suppression of these arrhythmias.
Tse et al working with an HCN1 mutant (HCN1-ΔΔΔ) with a deletion in the S3-S4 linker (position 235 to 237) created a biologic pacemaker in the procine atrium.21 The HCN1-ΔΔΔ mutation favors channel opening, and its expression in ventricular myocytes has been shown to result in automaticity with rates greater than 200 beats/min. In a porcine model of sick sinus syndrome, HCN1-ΔΔΔ was transduced with an adenoviral vector into the left atrium, and an electronic pacemaker was implanted. The HCN1-ΔΔΔ–injected pigs exhibited atrial pacemaking activity originating from the left atrium, which increased with catecholamines. The approach of Tse et al relies on normal atrioventricular (AV) nodal conduction to activate the ventricle. In a heart with impaired AV conduction, biologic pacemakers in the atrium will not effectively pace the ventricle.
Kv1.4 is a member of the Shaker K channel gene family. When expressed in heterologous systems, Kv1.4 channels express depolarization-activated delayed rectifier potassium currents. Furthermore, Kv1 gene family members are not expressed significantly in cardiac tissue. Kashiwakura et al created an ion channel based on Kv1.4, but functionally similar to HCN channels.22 This synthetic If-like channel was made via three point mutations within the voltage sensor in S4 (R447N, L448A, R453I): The channel was hyperpolarization activated and had a point mutation in the pore region (G528S), conferring permeability to K+ and Na+. Because the genes of Kv and HCN channels produce tetrameric ion channel complexes, hetero-multimerization with products from genes of the same family can occur. One advantage of a Kv1.4-based, If-like channel is that hetero-multimerization with native If channels (HCN genes) will not occur. A disadvantage of the synthetic Kv channel is that the cyclic adenosine monophosphate (cAMP) binding that is essential to the autonomic responsiveness of HCN channels is not replicated. Three to five days after injecting the synthetic Kv1.4 channel into guinea pig hearts, pacemaker function was detected. Patch clamp records of isolated myocytes revealed a robust hyperpolarization-activated inward current, and the myocytes also exhibited spontaneous action potentials.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


