CHAPTER 1 Anatomy of the Thorax
THORACIC CAGE
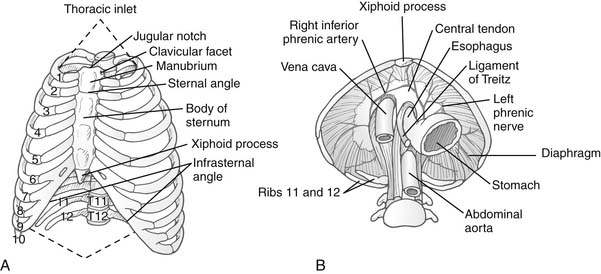
The bony thorax has two apertures, or openings: the superior thoracic aperture, often referred to as the thoracic inlet or cervicothoracic junction, and the inferior thoracic aperture (Fig. 1-1A).
Cervicoaxillary Canal
The cervicoaxillary canal is bounded by the first rib inferiorly, the clavicle superiorly, and the costoclavicular ligament medially (Fig. 1-2). The structures that pass through this space include the subclavian vein and artery and the brachial plexus.
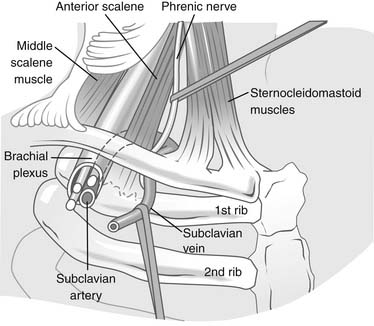
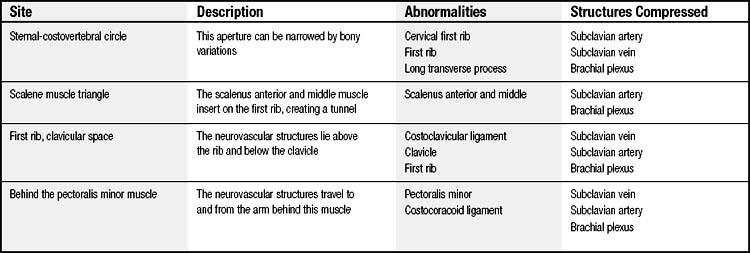
In the normal anatomy, there is ample room for each of these structures. There are four major areas in which the vessels or nerves can be compressed (Table 1-1).This topic will be discussed in greater detail in Chapter 26.
Diaphragm
The diaphragm has several openings through which structures can transverse from one cavity to another (see Fig. 1-1B). The inferior vena cava hiatus lies anteriorly and to the right of the midline, at the level of vertebra T8. The right phrenic nerve travels through this hiatus, and the left phrenic nerve penetrates laterally through its own opening at the same level. The esophageal aperture is at the level of the T10 vertebra, and the right and left vagal trunks that adhere to it enter the abdomen along with the esophagus.
The Bony Thoracic Cage
Ribs
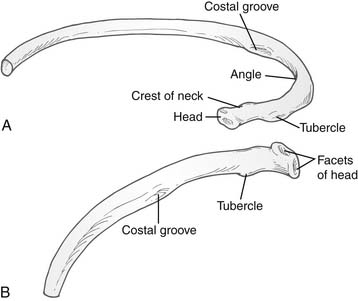
Ribs three to nine are classified as typical ribs and have a head, neck, and a shaft. The head has an upper and a lower articular facet divided by a crest for articulation with two adjacent vertebrae in synovial costovertebral joints, the lower facet articulating with the upper border of its own vertebra. The neck is flattened, with the upper border curling up into a prominent ridge—the crest. A tubercle projects posteriorly from the end of the neck and marks the junction of the neck and the body (Fig. 1-3).
MUSCLES OF THE THORAX
The muscles of the chest wall serve to protect the contents of the thoracic cavity, and they assist the movements of the thorax and upper extremities (Fig. 1-4). The 17 muscles of the chest wall are not discussed in detail here, but Table 1-2 lists them with their innervations and sites of attachments. Here we focus on muscle groups that are used in chest wall reconstruction. The latissimus dorsi, pectoralis major, serratus anterior, trapezius, rectus abdominis, and external oblique are the six major muscles that are available for reconstruction (Table 1-3).
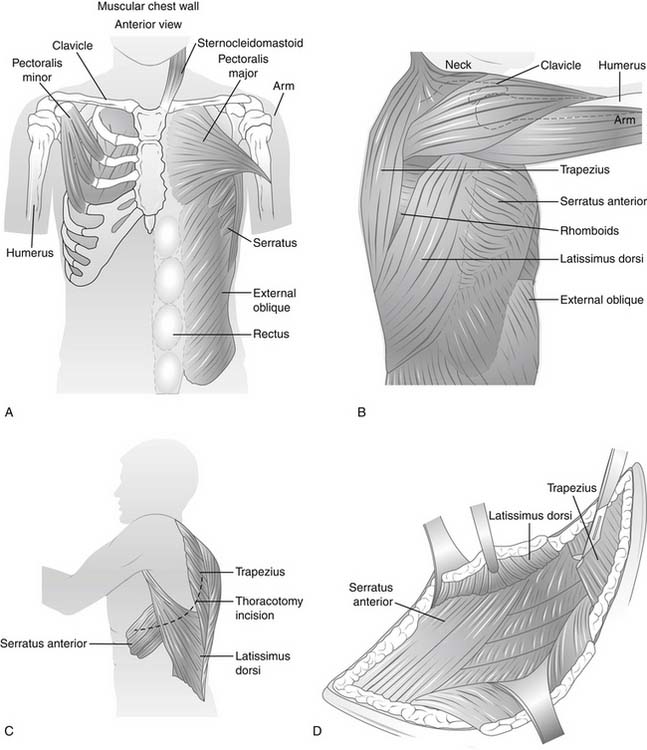
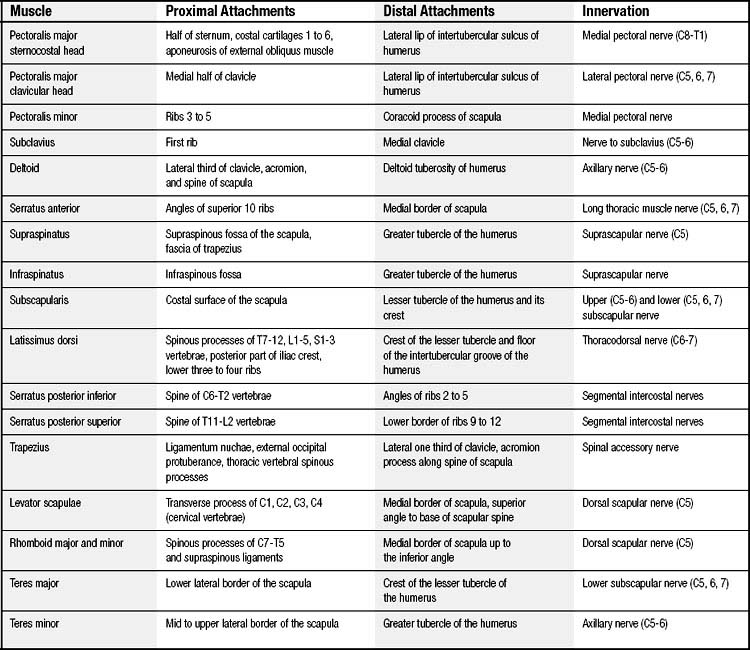
Table 1–3 Arterial Supply of Muscles Used in Chest Wall Reconstruction
| Muscle | Arterial Supply |
|---|---|
| Latissimus dorsi | Thoracodorsal artery |
| Pectoralis major | Thoracoacromial, lateral thoracic perforators, internal mammary artery, lateral intercostals artery |
| Serratus anterior | Thoracodorsal artery, long thoracic artery |
| Trapezius | Transverse cervical artery |
| External oblique | Lower thoracic intercostal arteries |
| Rectus abdominis | Superior and inferior epigastric arteries |
Intercostal Muscles
Fibers of the intercostalis externi muscle arise from the sharp lower border of the rib above and course inferomedially (i.e., in the direction of the fingers when the hands are put into the front pockets of trousers) to the smooth upper border of the rib below. Anteriorly, it is replaced by the anterior intercostal membrane. Between the bony ribs is muscle; between the costal cartilages is membrane. In the lower spaces, the muscle interdigitates with the fibers of the external oblique (Fig. 1-5).
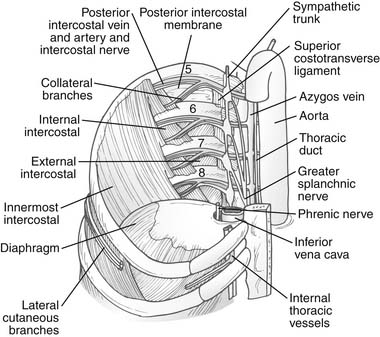
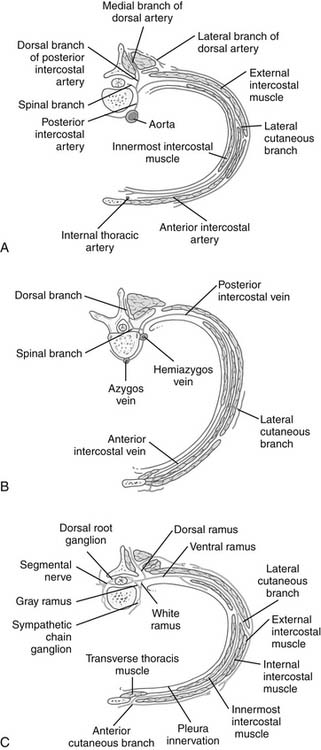
The intercostalis intimi muscles also traverse more than one space and are better developed in the lower lateral spaces. In the plane between the innermost and outer two layers runs the neurovascular bundle. From above downward, the order is vein, artery, and nerve (mnemonic: VAN). Beyond the angle of the rib posteriorly, they are protected by the downward projection of the lower border of the rib. Hence, for a thoracotomy, the periosteum is stripped off the upper half of the rib, avoiding the lower border and the neurovascular bundle (Fig. 1-6).
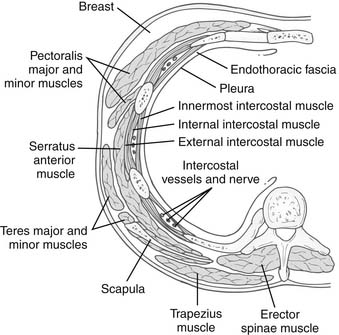
As in the rest of the body, the mixed spinal nerve is formed from a dorsal and a ventral root, the dorsal root being sensory and the ventral root containing somatic motor neurons. As it emerges from the intervertebral foramina, it branches into a dorsal and a ventral ramus. The dorsal ramus of the thoracic spinal nerve supplies the paravertebral back muscles and skin of the back. The ventral ramus communicates with the sympathetic chains via white rami communicantes (postganglionic fibers). Beyond this point, the true intercostal nerve lies just superficial to the parietal pleura in the endothoracic fascia. It gains the costal groove between the innermost intercostal and the internal intercostal muscles near the angle of the rib, where a collateral branch is given off. This small branch supplies the muscles of the space, the parietal pleura, and the periosteum of the rib. The main nerve itself has muscular branches, a lateral cutaneous branch, and a terminal anterior cutaneous branch. The lateral cutaneous branch given off along the midaxillary line gives off anterior and posterior branches to supply the skin over that space. Just lateral to the sternal margin, the terminal anterior cutaneous branches of the upper six nerves pierce the internal intercostal muscles, the external intercostal membrane, and the pectoralis major to reach skin. The lower five intercostal nerves slope downward behind the costal margin into the neurovascular plane of the abdominal wall. The subcostal or 12th thoracic nerve leaves the thorax by passing behind the lateral arcuate ligament and the subcostal artery and vein (Fig. 1-7). The cutaneous branches of each dermatome tend to overlap considerably; hence anesthesia after thoracic incisions is quite rare unless multiple intercostal nerves have been damaged. The first intercostal nerve is very small and supplies no skin, lacking both lateral and anterior cutaneous branches.
< div class='tao-gold-member'>
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


