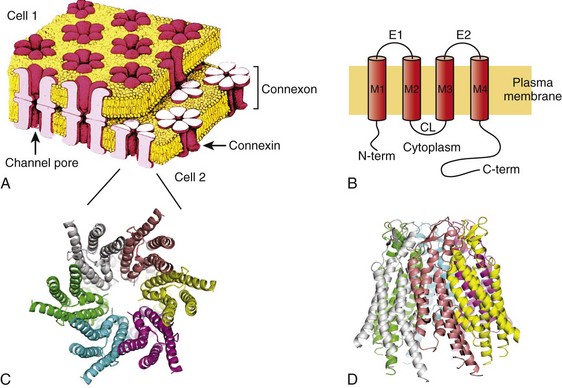
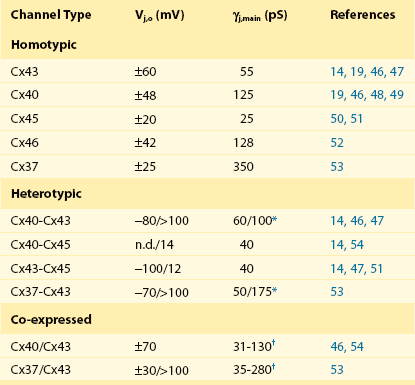
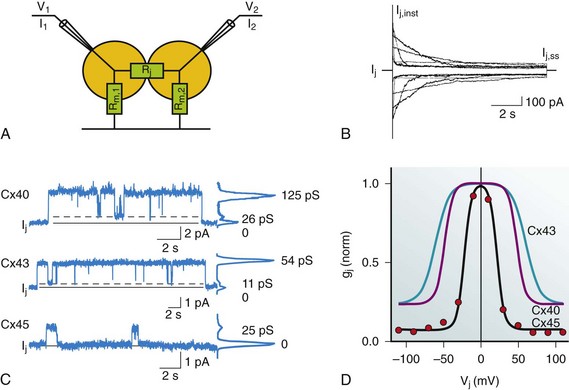
15 Structure of Gap Junction Channels Cardiac Gap Junctions: Homomeric, Heterotypic, and Heteromeric Forms Biophysical Properties of Cardiac Gap Junction Channels Action Potential Propagation in the Myocardium: The Role of Connexins Non–Voltage-Dependent Regulators of Channel Patency Extrinsic Uncoupling Agents of Cardiac Gap Junction Channels The propagation of the cardiac action potential throughout the working myocardium is made possible by voltage-dependent Na+, Ca2+, and K+ currents and by gap junction channels. One of the roles of gap junction channels is to permit the passage of currents from cell to cell that are essential for action potential propagation throughout the working myocardium. When considering action potential propagation gap junctions can be best understood as components of the longitudinal resistance within the functional syncytium of the myocardium. Gap junction channels are in series with the cytoplasmic resistance. The intercellular resistance of gap junctions is in series with the intracellular resistance of the cytoplasm, and together they represent longitudinal resistance. Both resistances can affect conduction velocity within the heart, but it is the gap junction channels composed of connexins that dominates longitudinal resistance. Dual whole-cell patch clamp studies of cardiac myocytes have been used to quantify gap junction membrane resistance or junctional conductance in vitro. The estimates of junctional resistance for ventricular myocyte cell pairs reveal that it is often an order of magnitude less than the input resistance of an isolated myocyte, somewhere in the range of 2 to 10 MΩ or 1000 to 100 nS.1 These values must be considered under estimates because of the series resistance of the pipettes that have similar resistance values to the junctions themselves.1 In addition to creating an electrical syncytium for the heart gap junctions also allow the passage of small solutes such as cAMP that are able to affect the function of multiple systems within cardiac myocytes. To better understand how gap junction channels contribute to the normal functions of the heart and how they participate in cardiac arrhythmias and ischemia, it is necessary to first describe their structure and biophysical properties. Structural analysis has revealed that gap junction channels are composed of two hemichannels or connexons linked together that provide an intercellular pathway between two adjacent cell interiors (Figure 15-1, A). Each hemichannel is composed of six connexins. Each connexin contains four membrane-spanning domains (M1-M4; see Figure 15-1, B). The N-terminus protrudes from M1 into the cytosol. M1 and M2 are connected by an extracellular loop E1, whereas M2 and M3 are connected by a cytoplasmic loop. M3 and M4 are connected by another extracellular loop (E2), and the cytoplasmic extension of M4 is the C-terminus (see Figure 15-1, B). This general model is true for all the cardiac connexins and the other human connexins. Individual gap junction channels within the atrial and ventricular myocardium are found in abundance at the intercalated discs in the form of membrane complexes or plaques containing a large number of channels, ranging from hundreds to thousands. A small portion of such a plaque is depicted in Figure 15-1, A. These plaques can also form along the lateral surfaces of myocytes in normal myocardium and can become even larger structures in stressed or diseased myocardium.2 Figure 15-1 A, A portion of a junctional plaque. Yellow bilayers are the plasma membranes of two adjacent cells. Each gap junction channel is composed of two hemichannels or connexons that are themselves composed of connexins. Modified after reference.55 B, Schematic representation of a connexin with 4 membrane domains, M1-M4, 2 extracellular loops, E1 and E2, and 3 cytoplasmic domains, N-terminus, cytoplasmic loop, and the C-terminus. C, En face view the hemichannel from the cytoplasm of a cell. D, Molecular rendition of connexin26 (Cx26) viewed from within the plasma membrane of a cell. Each connexin is colored differently. (A, Modified from Makowski L, Caspar DL, Phillips WC, et al: Gap junction structures. VI. Variation and conservation in connexon conformation and packing. Biophys J 45:208–218, 1984.) Only a few studies have been successful in elucidating the molecular structure of gap junction channels using electron crystallography. The first analysis was performed on a noncardiac connexin, Cx26, and more recently has been revisited, revealing structural detail to a resolution of approximately 0.35 nm.3 Cx43 has also been analyzed using crystallographic methods with a resolution of approximately 1.0 nm.4 Figure 15-1, C, depicts a representation of the helical membrane domains and the extracellular loops of a hemichannel as viewed from within the plasma membrane of a cell. Figure 15-1, D, illustrates the structure of Cx26 from the perspective of looking down its long axis directly into the channel from the cytoplasm where the four transmembrane domains of each connexin can be visualized. The structural analysis has not been sufficiently detailed to demonstrate clearly which membrane-spanning domains are forming the channel wall. However, the structure shown in Figure 15-1, D, again depicting the four transmembrane helices, suggests that M1, M2, M3, and even M4 are potential contributors to the lining of the pore or the cytoplasmic vestibule. One approach that has been used to define the pore lining regions of connexins has been the substituted cysteine accessibility method (SCAM). Substituting cysteine for other amino acids thought to be contributing to the pore wall is the first step to successful use of SCAM. This step is followed by a demonstration that the substitution does not affect normal channel activity. To establish whether the substituted group is part of the pore wall, a thiol reactive agent such as maleimide or a derivative is then perfused into the preparation while monitoring channel activity. One possible outcome is altered unitary conductance consistent, with the substituted group being a component of the pore wall. In fact, the application of SCAM to connexins has provided varied results when assessing pore lining regions of the four membrane-spanning domains and extracellular loops and intracellular regions. The use of SCAM to elucidate the pore wall structure on Cx46 has generated data most consistent with pore lining domains or pore access regions in M1 and the E1 loop,5 but Cx32 SCAM data are more consistent with M3 as a major pore lining helix.5 No SCAM analysis has been performed on Cx43, Cx40, Cx45, or Cx37. Site-directed mutagenesis has also been used for Cx43 and Cx37 with various substitution strategies. In Cx43, mutations were introduced in M3 that resulted in silent channels.5 This is not conclusive evidence that M3 participates as a functional member of the pore wall, but it does not exclude the possibility. For Cx37, site-directed mutations in M3 resulted in altered conductive states, which is consistent with M3 participation in forming the pore wall, but does not exclusively demonstrate that either.6 The crystallographic and mutagenic data lead to one possible explanation: different connexins do not use the identical structural motifs in the formation of an intercellular pore despite functional similarities. The crystallographic structure shown in Figure 15-1, C, indicates that there are potential surfaces for all four domains contributing to the channel or pore and the cytoplasmic vestibule. Hemichannels composed of six connexins from two closed aligned cells form a linkage via the extracellular domains E1 and E2 to create a complete gap junction channel. The heart does not express all 21 identified connexins7 that are able to assemble into functional gap junction channels (see Figure 15-1, A, B). Instead, a select number of connexins are expressed within the human heart; they are Cx43, Cx40, Cx45, Cx46, and Cx37.8 Their distribution within the heart is not uniform. For example, Cx43 is abundantly expressed within the ventricles but is only sparingly expressed within the AV and SA nodes.8,9 Table 15-1 illustrates the relative connexin expression levels within the ventricles, atrium, SA node, AV node, and bundle branch/conducting system (BB/CS). Connexin 31.9, the ortholog of mouse Cx30.2 found in the mouse AV node, has thus far not been detected in the human heart. Cx30.2 has been shown to be in part responsible for conduction delay at the AV node in mice.9 The absence of Cx31.9 in the human AV node suggests that the delay in humans might be the result of reduced channel numbers or the presence of heteromeric or heterotypic forms of Cx45, Cx40, and Cx43 or possibly the existence of another unidentified connexin within the AV node whose properties mimic those of mouse Cx30.2. Table 15-1 BB/CS, Bundle branch/conducting system; AV, ventricular; SA, sinoatrial. From Severs NJ, Bruce AF, Dupont E, et al: Remodelling of gap junctions and connexin expression in diseased myocardium. Cardiovasc Res 80:9–19, 2008; and Jansen JA, van Veen TA, de Bakker JM, et al: Cardiac connexins and impulse propagation. J Mol Cell Cardiol 48:76–82, 2010. As implied in Table 15-1, myocytes within the different regions of the heart are able to coexpress connexins. For example, individual atrial myocytes express Cx40, Cx43, and Cx45 simultaneously but in differing amounts with Cx40 being the most abundantly expressed connexin. The expression of a single connexin within myocytes has the potential to generate functional gap junction channels composed of two identical hemichannels, both composed of the same connexin referred to as homomeric (i.e., homotypic) channel. Another type of gap junction channel is also possible, where each hemichannel of two opposing cells is homomeric but each cell expresses a different connexin. This type of channel is heterotypic.7 Finally, because most myocytes express at least two and often three connexins, a hemichannel can potentially contain two or possible three different connexins.7,10 This type of hemichannel is heteromeric. Two heteromeric hemichannels will form a heteromeric gap junction channel. The biophysical properties of gap junction channels are best illustrated using a dual whole-cell patch clamp on isolated cell pairs. Figure 15-2, A, depicts a cell pair coupled by gap junctions with the equivalent circuit for the cell pair. All the cardiac gap junction channels have been studied in connexin-deficient cells that are then transfected with specific cardiac connexins to better understand how homotypic gap channel forms of Cx43, Cx40, Cx45, Cx46, and Cx37 behave. In all cases, each can be shown to gate closed with the application of increased transjunctional voltage (Vj). A macroscopic record of junctional currents is shown in Figure 15-2, B, for Cx45 and illustrates the decline of junctional current with sustained Vj. Ij,inst is the junctional current recorded at the onset of a voltage step, and Ij,ss is the steady state current. In some cases, individual channel activity can be observed as shown in Figure 15-2, C. Single-channel recordings for Cx43, Cx40, and Cx45 are shown. Multichannel and single-channel data have allowed the determination of unitary conductance (γj,main) for the cardiac connexins, which are listed in Table 15-2. The ability to monitor unitary events has also allowed a better understanding of voltage-dependent gating in connexins, which has been shown to have at least two distinct mechanisms: fast gating and slow gating. Fast gating is characterized by a rapid transition from an open state to a residual state (γj,residual; see dashed lines in Figure 15-2, C) or closed state, whereas slow gating is manifest as a series of subconductive states transitioning from an open or closed state.10 A systematic study on Cx30 gap junctions reported five substrate conductances unevenly spaced between γj,main and γj,residual, suggesting that each of the six connexins of a hemichannel act as a subgate.11 Macroscopic recordings of junctional currents (see Figure 15-2, B) have also been useful in dissecting the molecular mechanisms of voltage-dependent gating. Plotting the steady state currents generated in response to different Vj steps (see Figure 15-2, B) reveals the relationship between junctional conductance (gj) and transjunctional potential (Vj). gj is derived from the ratio of gj,ss/gj,inst. A Boltzmann fit of the normalized steady state junctional conductance for various amplitude voltage steps is shown in Figure 15-2, D, for Cx43, Cx40, and Cx45. Table 15-2 Multichannel and Single-Channel Data of Different Types of Gap junction Channels *Conductance polarity dependent. †Conductances from homotypic, heterotypic, and heteromeric channels are possible. Figure 15-2 A, Dual whole-cell patch clamp equivalent circuit and superimposed cell pair. The patch electrodes ideally are significantly lower resistance than the junctional (Rj) and nonjunctional resistances (Rm). The circuit allows for the computation of junctional conductance (gj) with the following relationships: Vj = V2 − V1; I1 = Im,1 + Ij; I2 = –Ij; gj = Ij / Vj. B, Macroscopic record of homotypic Cx45 using Vj steps of ±10, ±30, ±50, ±70, ±90, and ±110 mV of 10 seconds in duration, which illustrates the voltage dependence of Cx45 and the varied time constants of inactivation with Vj. Ij,inst is the instantaneous junctional current and Ij,ss is the steady state current. C, Single-channel properties of Cx43, Cx40, and Cx45 channels. Records of single-channel currents observed in pairs of HeLa cells transfected with Cx40, Cx43, and Cx45. Solid line, zero current level; dashed line, residual current level. The current histograms (right) revealed γj,main = 125 pS and γj,residual = 26 pS for Cx40, γj,main = 54 pS and γj,residual = 11 pS for Cx43, and γj,main =24 pS for Cx45 cell pairs; pipette was filled with 120 mM K+aspartate–. D, Normalized gj plotted against Vj for Cx43, Cx40, and Cx45. The red dots represent the record shown in (B). Normalized junctional conductance (gj) is steady state conductance gj,ss normalized against the instantaneous conductance gj,inst. (Data modified from Valiunas V, Beyer EC, Brink PR: Cardiac gap junction channels show quantitative differences in selectivity. Circ Res 91:104–111, 2002; and Valiunas V, Gemel J, Brink PR, et al: Gap junction channels formed by coexpressed connexin40 and connexin43. Am J Physiol Heart Circ Physiol 281:H1675–H1689, 2001.) An important parameter derived from the Boltzmann fit is Vj,o. This parameter represents the half inactivation voltage or that transjunctional potential where half the channels can be considered closed. The instantaneous junctional conductance remains relatively constant regardless of the voltage, implying that many if not all the gap junctions are patent when Vj is zero. Analysis of single and multichannel records of Cx43 (see Figure 15-2, C) have been used to determine open probability under chronic application of Vj between 20 and 40 mV yielding values ranging from 0.5 to 0.95, consistent with the idea that gap junction channels are in the open state when there is no applied transjunctional potential. The application of Vj greater than 50 mV reduces the mean open time, whereas mean closed time remains relatively constant, which translates into reduced open probability (Po) with increased Vj amplitude.12 This is consistent with the behavior manifest in macroscopic recordings for Cx43. Nonstationary analyses of Cx43 and Cx37 have also revealed similar results with open probabilities less than 0.95 for Cx43 and 0.7 for Cx37.13 Table 15-2 lists Vj,o and unitary conductance for the cardiac connexins, which includes homotypic, heterotypic, and heteromeric forms. The values of Vj,o for homotypic channels vary from 20 mV for Cx45 to 60 mV for Cx43. Also listed are all the Vj,o values for heterotypic forms that have thus far been determined. In addition, values for cells coexpressing Cx43/Cx40 and Cx43/Cx37 are given. Unitary conductance for the cardiac connexins varies greatly, as seen in Table 15-2. For heterotypic forms, the observed unitary conductance can be polarity dependent.14 Observation of cell pairs coexpressing connexins reveals a range of conductances consistent with channels of different connexin content. The macroscopic junctional currents shown in Figure 15-2, B, illustrate that the currents decline (deactivation) with a prescribed time course on the order of 0.1 to 1 seconds for Cx43.15 The time constant for voltage-dependent inactivation is itself voltage dependent and becomes shorter with larger transjunctional voltage steps. The time course of voltage-dependent inactivation in junctional currents as shown in Figure 15-2, B, for Cx45 are similar to the behavior of the other cardiac connexins, although it should be noted that Cx45 has the largest inactivation time constants of the cardiac connexins, approximately 0.1 seconds for large transjunctional potentials and 3 seconds for smaller transjunctional steps.15
Biophysical Properties of Gap Junctions
Background
Structure of Gap Junction Channels
Cardiac Gap Junctions: Homomeric, Heterotypic, and Heteromeric Forms
Ventricles
Cx43 > Cx45 > > Cx40 > > Cx46~Cx37
Atrium
Cx40 > Cx43 ~ Cx45
BB/CS
Cx40 ~ Cx43 ~ Cx45
AV node
Cx45 > Cx40 ~ Cx43
SA node
Cx45 > Cx40 > #x003E; #x003E; > Cx43
Biophysical Properties of Cardiac Gap Junction Channels
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Thoracic Key
Fastest Thoracic Insight Engine
