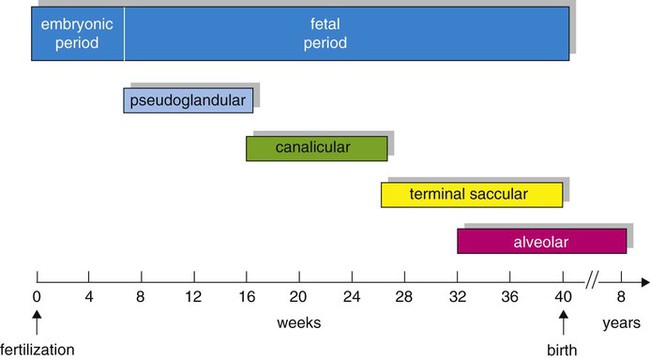
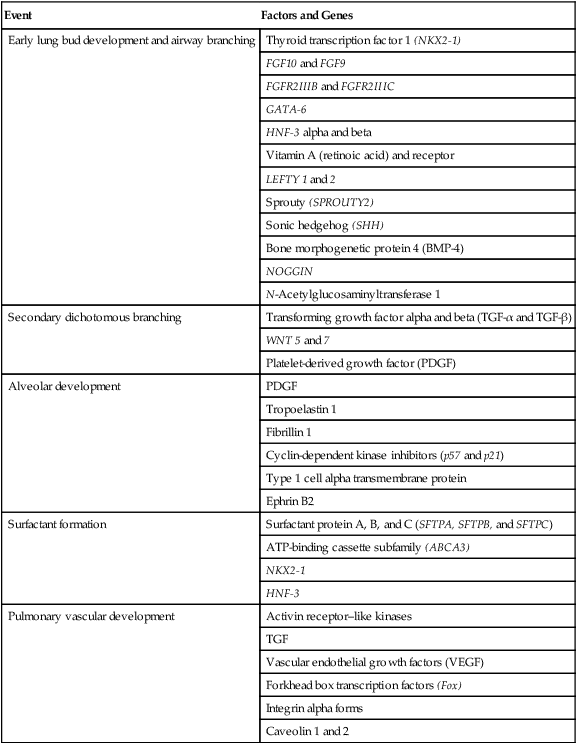
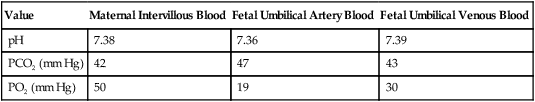
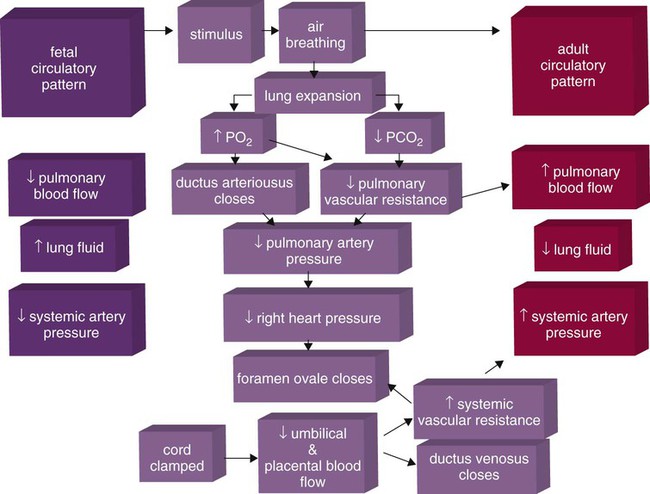
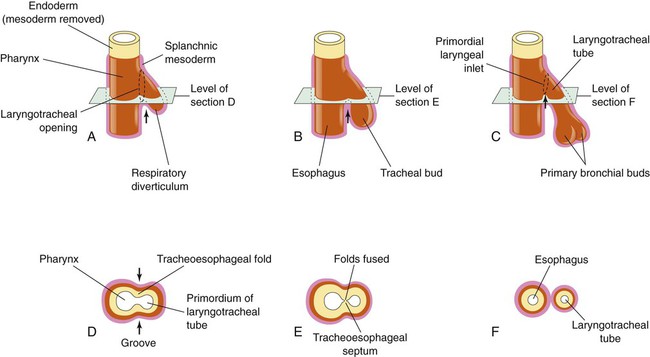
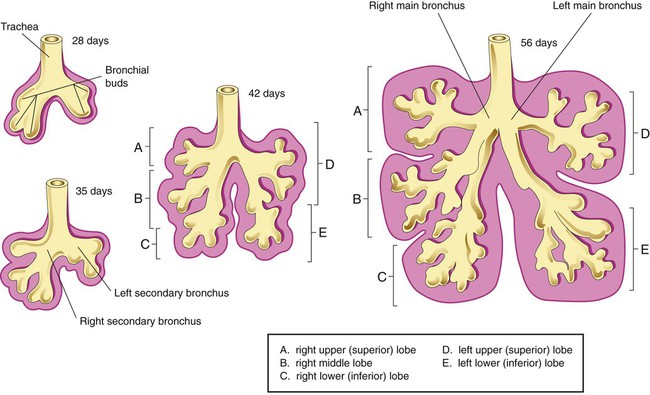
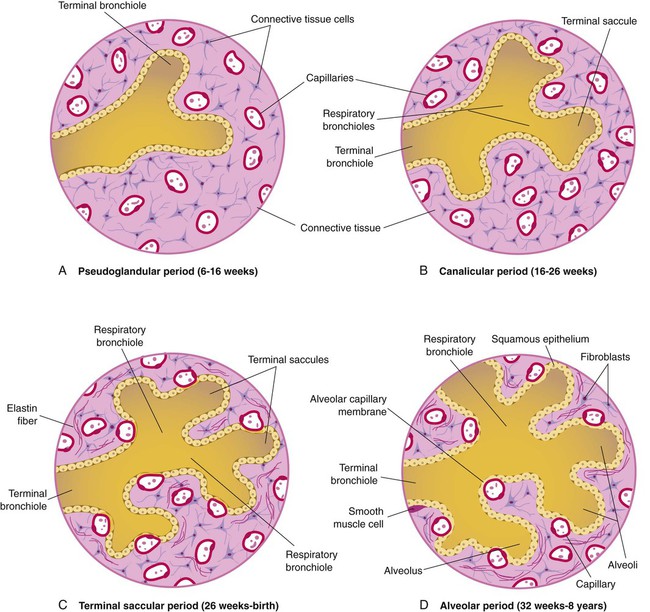
After reading this chapter you will be able to: Based on cellular differentiation and tissue architecture, development of the respiratory system has been categorized into various stages.1 Figure 8-1 shows the various stages of lung development, and Table 8-1 summarizes the major developmental events in each phase. Respiratory development begins in the embryonic period on or about day 22 after fertilization, when a small mass of cells, the respiratory primordium, begins to develop near the ventral region of the fourth pharyngeal arch of the primitive pharynx. This mass of cells forms a pouchlike bud, the respiratory diverticulum, on about day 26 (Carnegie stage 9) that continues to grow to form a laryngotracheal tube (Figure 8-2). The laryngotracheal tube forms from a groove in the fourth pharyngeal pouch. From the laryngotracheal tube, a tracheal bud forms by the end of the fourth week of life. The dorsal portion of the primitive foregut develops into the primordial esophagus and is separated from the tracheal bud by the formation of a tracheoesophageal septum. During week 5 of development, the tracheal bud continues to develop and bifurcates into left and right primary bronchial buds. The laryngeal structures develop at the superior end of the laryngotracheal bud. TABLE 8-1 Developmental Events of the Cardiopulmonary System The developmental branching process of the airways and blood vessels of the lung is highly regulated by the timely activation of various genes in different locations. Of the approximate 22,000 genes in the human genome, about 40 are required for normal respiratory development.2–4 Table 8-2 lists many of these genes and the process in which they play an important role. The initial step in the development of the respiratory system is the localized expression of the NKX2-1 gene (also known as thyroid-specific transcription factor, TTF-1) in the anterior wall of the foregut, which stimulates the primary lung bud formation. Failure or mutation of the NKX2-1 gene can lead to failure of lung bud formation and various tracheoesophageal malformations.5 Lung bud elongation and the repetitive airway branching process is stimulated and directed by the highly choreographed expression of other key genes, including FGF10, FGFR2IIB, GATA-6, HNF-3, SPROUTY2, SHH, BMP-4, and NOGGIN as well as numerous other genes (see Table 8-2). Mutations of the FGF10 gene can result in tracheal development but fatal failure of further lung formation.6 TABLE 8-2 Genes Implicated in Pulmonary Development At approximately 6 weeks of development, lung and airway growth has the appearance of a glandular structure—hence the name of the second phase of development, the pseudoglandular stage (Figure 8-3). For the next 10 weeks, the growth and branching of the tracheobronchial tree and pulmonary vasculature continue, under the direction of the various genes described earlier, and culminate with formation of the terminal and respiratory bronchioles. The distinction between these two types of bronchioles is important. Terminal bronchioles, similar to bronchi and the trachea, are conducting airways only and do not participate in gas exchange with blood. Respiratory bronchioles have much more superficial capillaries and are capable of gas exchange with blood and become more elaborate as development continues. Branching and dividing of the tracheobronchial tree occur in several ways as the result of differential gene expression. A single bud that develops off of an existing structure is termed a monopodial bud. Airways that divide into two or more airways do so through dichotomous branching. Most of the divisions of airways occur in a nonsymmetric fashion termed irregular dichotomous branching.7,8 The epithelial lining of the airways begins to differentiate into columnar epithelia in the proximal airways and differentiates into cuboidal epithelium in the more distal bronchioles (Figure 8-4, A). Development of cilia, mucous glands, and goblet cells occurs at this time, and these are found lining most of the conducting airways. The third phase of development is termed the canalicular stage (see Figure 8-4, B). It begins at week 16 and continues until week 26. The canalicular stage overlaps with the pseudoglandular stage because the superior regions of the lung are developing slightly faster than the inferior regions. During this phase, primary changes include the development of two to four more generations of respiratory bronchioles from each terminal bronchiole, the formation of blind tubular alveolar ducts from each respiratory bronchiole, and greater blood vessel development. In the last several weeks of this stage, the region beyond each terminal bronchiole forms the functional structure called the acinus, the basic gas-exchanging unit of the lung. At this time, the two principal epithelial cell types that cover the gas exchange surface begin to appear, type I and type II pneumocytes. At the end of the canalicular period (24 to 26 weeks of gestation), the fetus, if born, is capable of sufficient gas exchange and is viable if supported with supplemental O2, ventilatory support, and surfactant administration. During the fourth phase, the terminal saccular stage (see Figure 8-4, C), more terminal bronchioles and their associated acini form, and their structure continues to develop from 26 weeks to birth. The formation of the total number of terminal bronchioles is complete at the end of this phase.8 The cuboidal epithelia that line the blind tubules of the acinus continue to differentiate into rounded secretory cells (type II pneumocytes) and flatter squamous epithelial cells (type I pneumocytes). Mounting evidence shows that an important source of type I pneumocytes during both development and after lung injury are type II cells that can proliferate and differentiate.9,10 Capillaries continue to form near and bulge from the surface of the acinus. Although some type II pneumocytes form by 20 weeks’ gestation, they are in such small numbers and of such primitive function that their impact on lung function is marginal. From this point until birth, there is rapid proliferation of alveolar ducts and sacs, formed from the respiratory bronchioles. The type I pneumocytes of the saccule walls thin and elongate to cover the walls of this region. Type I cells become the primary gas-exchange cells in the lung with close approximation to developing pulmonary capillaries. Type II pneumocytes form and secrete the vital pulmonary surfactants that are necessary to alter surface tension and help keep the lungs inflated. The development of mature alveoli, accompanied by capillary proliferation within the walls, marks the final phase of lung development and is known as the alveolar period (see Figure 8-4, D). This phase begins at about week 32 of gestation and continues for years after birth. During this phase, the terminal saccules develop pouchlike regions called alveoli in their walls that are hexagonal in shape. The process of alveolarization occurs through the formation of crests along the immature airway wall, which develop further into septa that lengthen into the terminal saccule lumen; this effectively divides up the terminal airspace and results in greater numbers of alveoli that enlarge to a mature state with time. A full-term newborn infant has about 50 million alveoli, and the number continues to increase for about 2 to 3 years after birth.11,12 The alveoli are lined with type I and II pneumocytes covering the pulmonary capillaries that have formed just below the basement membrane. Human pulmonary surfactant, which promotes lung inflation and protects the alveolar surface, begins to be produced around 24 to 25 weeks of development by type II pneumocytes. It is composed primarily of phospholipids, a small amount of protein (types SP-A, SP-B, and SP-C), and a trace of carbohydrates.13 Early research in pulmonary surfactants centered on the phospholipid components, mainly phosphatidylcholine (lecithin [L] and sphingomyelin [S]) and phosphatidylglycerol (PG). Quantification of these phospholipids (the L/S ratio and PG concentration) provides a predictive index of the lung maturity in a fetus before birth and the risks of the development of respiratory distress.14 An L/S ratio of 2 or more indicates a relatively low risk for the development of respiratory distress syndrome, whereas an L/S ratio of less than 1.5 is associated with a high risk. Surfactant synthesis is regulated by numerous hormones and factors, including glucocorticoids, prolactin, insulin, estrogens, androgens, thyroid hormones, and catecholamines.15 Glucorticosteroid production increases at the end of gestation and stimulates receptors in type II pneumocytes to increase surfactant production and improve the L/S ratio. Various key genes are also associated with normal surfactant production (surfactant protein genes A, B, C, and D; surfactant protein A, B, C, and D; and an adenosine triphosphate (ATP)–binding cassette transporter, ABCA3), and their failure, owing to mutation, is linked with the development of respiratory distress syndrome and other pulmonary disorders.16 A distinctive function of the developing lung is the formation of relatively large amounts of fetal lung fluid that is passed into amniotic fluid. Fetal lung fluid is a unique combination of plasma ultrafiltrate from the fetal pulmonary microcirculation, components of pulmonary surfactant, and other fluids from pulmonary epithelial cells.7 This fluid is constantly produced and keeps the fetal lung inflated at a slight positive pressure with respect to amniotic fluid pressure; it is important in stimulating normal lung development.17 At term, the fetal lung is filled with about 40 ml, and fluid is produced at a rate that results in replacing it multiple times per day. Conditions that lead to reduced fetal breathing and amniotic fluid formation (oligohydramnios) are linked to incomplete inflation of the lung with fluid and poorly developed (hypoplastic) lungs. Throughout the developmental period, lung growth is similar in male and female fetuses. There are differences, however. At birth, the lungs of male infants are, on average, larger and have a greater number of respiratory bronchioles than the lungs of female infants when adjusted for gestational age.18 When evaluating breathing efforts and surfactant production at 26 to 36 weeks of gestation, female fetuses have better developed lung function and are slightly less susceptible to the development of respiratory distress syndrome.19,20 Survival of the embryo and then fetus requires an effective circulatory interface with the circulation of the mother, which is provided by the placenta.21 Within 1 week of uterine implantation, vascular projections called chorionic villi arise from the aorta of the embryo and penetrate the uterine endometrium. As gestation proceeds, the villi increase in number and complexity; erode the endometrium; and create irregular pockets called intervillous spaces in the placenta, which fill with maternal blood. The maternal blood flowing through the intervillous spaces bathes the embryonic villi and creates an O2-rich and nutrient-rich blood environment. As gestation progresses, the villi decrease in size but increase in number and complexity, resulting in an increased surface area that is essential for adequate maternal-fetal gas, nutrient, and waste exchange. The maternal uterine tissues and blood vessels of the fetal chorionic villi make up the bulk of the placenta. Figure 8-5 shows a cross section of a well-developed placenta. Maternal blood flows into the intervillous space through the spiral arteries, whereas fetal blood is supplied to the villi from two umbilical arteries. Maternal and fetal blood come into close proximity but remain separated by an embryonic membrane that permits the exchange of O2, CO2, water, ions, various metabolic molecules, and hormones. Some maternal cells do move into fetal blood, and some fetal cells move into maternal blood and have been found in various maternal organs. Various factors enhance the delivery of O2 to fetal tissues. The partial pressure gradient for O2 between maternal blood and fetal blood drives the diffusion of O2 into fetal blood within the chorionic villi capillaries.22,23 The maternal arterial blood has a partial pressure of O2 (PaO2) of approximately 100 mm Hg, which mixes with the blood in the intervillous space to produce a mean PO2 of approximately 50 mm Hg. Fetal blood that enters the villi has a PO2 of approximately 19 mm Hg, and the pressure gradient between maternal and fetal blood PO2 (50 − 19 = 31 mm Hg) causes O2 to diffuse into fetal blood. Blood leaving the villi and entering the umbilical vein has a PO2 of approximately 30 mm Hg. Table 8-3 summarizes the normal gas and acid-base values in normal fetal umbilical arteries and veins and maternal intervillous blood. Assessment of umbilical vein blood gas data shortly after birth is a method of determining the degree of fetal asphyxiation during the birth process. TABLE 8-3 Approximate Normal Values of Blood Gases and Acid-Base in Fetal and Maternal Blood The O2 content and delivery by fetal blood are almost the same as adult blood despite the much lower PO2; this is due to several factors, including relatively higher content of hemoglobin (18 g/dl) and hematocrit (54%) in fetal blood and the presence of fetal hemoglobin (HbF), which has an increased affinity for O2 and a more pronounced Bohr effect (reduced oxyhemoglobin affinity with acidosis) to enhance O2 release.23 Figure 8-6 illustrates how the increased O2 affinity is manifested by a leftward shift of the fetal oxyhemoglobin dissociation curve. The P50 (PO2 that saturates 50% of the hemoglobin) is 6 to 8 mm Hg less than the P50 for adult hemoglobin (HbA), which indicates the degree of the shift toward higher affinity. At birth, approximately 70% of circulating hemoglobin is HbF. HbA gradually replaces HbF during the first 6 months of extrauterine life as HbA genes in bone marrow switch on and HbF genes in the liver (major site of fetal erythrocyte development) are switched off. Fetal circulation is different than the circulation of the neonate after birth.24 Three important bypass pathways function in the developing fetus to enhance the flow of blood to the developing organs: ductus venosus, ductus arteriosus, and foramen ovale. Oxygenated blood from the placenta is carried in the umbilical vein back to the fetal circulation via the hepatic circulatory system (Figure 8-7). Approximately one-third of this blood flows to the lower trunk and extremities. The other two-thirds flows through the ductus venosus, which bypasses the liver’s circulation and flows to the inferior vena cava. This better oxygenated blood in the inferior vena cava mixes with the venous blood returning from the lower trunk and extremities and enters the right atrium. Approximately 50% of this blood is shunted from the right atrium into the left atrium through an opening in the interatrial septum called the foramen ovale. Left atrial blood flows to the left ventricle and then to the ascending aorta, where it continues on to the brain, brachiocephalic trunk, and descending aorta. Venous blood from the superior vena cava is directed downward through the right atrium into the right ventricle and then into the main pulmonary artery. Various mechanisms work together to reduce and clear the amount of lung fluid at birth in preparation for air inflation.25 Days before birth, the epithelia of the lung stop the production of lung fluid. The lung fluid is actively absorbed back into the fetal circulation. Most of the active lung water absorption is facilitated by active sodium channel activity that is stimulated by fetal and maternal thyroid hormones, glucocorticoids, and epinephrine and increasing fetal lung and blood O2 content. In addition, some evidence suggests that the water channel aquaporin is also active in this process.26 During normal vaginal delivery, approximately one-third of the lung fluid is cleared through compression of the thorax in the birth canal. The pulmonary capillaries and lymphatics clear the remaining fluid. The newborn infant is stimulated by new tactile and thermal stimuli, all of which stimulate breathing. In addition, as placental gas transfer is suddenly interrupted, the newborn quickly becomes hypoxemic, hypercapnic, and acidotic. This situation triggers strong inspiratory efforts (Figure 8-8). At first, no air enters the newborn lung until the transpulmonary pressure gradient exceeds 40 cm H2O. As lung volume increases in a stepwise fashion with each breath, increasingly less pressure is needed to overcome the opposing forces. The volume trapped in the lung stabilizes quickly and is crucial to adequate gas exchange. Figure 8-9 summarizes the major cardiopulmonary changes that occur during the transition from a fluid-filled lung to an air-filled lung. As the lung expands with air, and gas exchange starts within the lung, pulmonary blood PO2 increases, PCO2 decreases, and pH increases; this results in pulmonary vasodilation, lower pulmonary vascular resistance, and constriction of the ductus arteriosus, which facilitates greater blood flow through the pulmonary circulation. Ductus arteriosus closure is stimulated further by the loss of maternal prostaglandins. The combination of increasing alveolar air content and constriction of the ductus arteriosus promotes progressive improvement in the matching of ventilation and blood flow, which increases the PO2 and decreases the PCO2 of blood leaving the lungs. After the clamping of the umbilical cord, cessation of umbilical and placental blood flow causes closure of the ductus venosus and a rapid increase in systemic vascular resistance. As systemic vascular resistance increases, left-sided heart pressures increase. Left atrial pressures also increase as a result of increased pulmonary blood flow that returns from the lungs. With left-sided heart pressures now higher than right-sided pressures, the foramen ovale closes. Figure 8-10 shows the relative differences of the upper airway in relation to body size in an infant and an adult. The greater relative weight of the head can cause acute flexion of the cervical spine in infants with poor muscle tone. Infant neck flexion causes acute airway obstruction. Although the head is larger, an infant’s nasal passages are proportionately smaller than those of an adult. In addition, the infant’s jaw is much rounder, and the tongue is much larger relative to the size of the oral cavity.27 These anatomic differences increase the likelihood of airway obstruction when an infant becomes unconscious and loses muscle tone. Most infants breathe through the nose. However, most term newborn infants shift to oral breathing in response to nasal occlusion and hypoxia.28 As normal infants mature, they begin to use the oral breathing route more and are more capable of shifting to oral breathing when nasal obstruction is present.29 At approximately 4 to 5 months of age, most infants are capable of full oral ventilation. A newborn’s larynx lies higher in the neck compared with the larynx of an adult, with the glottis located between C3 and C4, and is more funnel-shaped than that of an adult. In a child, the narrowest region of the upper airway is through the cricoid cartilage, rather than the glottis, as it is in adults. The epiglottis of an infant is longer and less flexible than the epiglottis of an adult and lies higher and in a more horizontal position. During swallowing, the infant’s larynx provides a direct connection to the nasopharynx. This connection creates two nearly separate pathways, one for breathing and one for swallowing, allowing infants to breathe and suckle at the same time. Anatomic descent of the epiglottis begins at The large conducting airways of infants are shorter and narrower than the airways of adults. The normal newborn trachea is approximately 5 to 6 cm long and 4 mm in diameter, whereas in small preterm infants, it may be only 2 cm long and 2 to 3 mm wide. Because of the smaller airways, a newborn’s anatomic dead space is proportionately smaller than the anatomic dead space of an adult, being approximately 1.5 ml/kg of body weight. Figure 8-11 compares the tracheal anatomy in an adult and a newborn. The main stem bronchi branch off from the trachea in the infant at less acute angles than in the adult. However, similar to adults, the right main stem bronchus of the infant is still more in line with the trachea, which promotes right main stem intubation when airways or suction catheters are inserted to deeply. Mean airway diameter, from main bronchi to respiratory bronchioles, increases about two to three times from birth to adulthood.31 The human lung continues to develop alveoli for years until it reaches a stable stage, at which the total number has increased to approximately 480 million alveoli.32 All development is generally complete by 10 years of age with most occurring in the first It was previously thought that the above-described alveolar development process ended several years after birth. However, numerous studies in various mammals have shown that compensatory lung growth can rapidly occur in the lung when part or all of the other lung is removed.36–38 Stem cell activation in the lungs, in response to gene and mechanical stretch, appears to be responsible for alveolar development well into adulthood after loss of lung tissue.39 The basic architecture of the pulmonary circulation is complete at birth. The main pulmonary trunk arises from the right ventricle and divides into left and right pulmonary arteries that supply each lung. These arteries divide further to form direct or conventional arteries and supernumerary arteries. Conventional arteries follow the airway branching, whereas supernumerary arteries follow an irregular pattern that allows substantial collateralization of flow between different regions of lung. Both types of pulmonary arteries come together to supply blood to large clusters of alveoli that are supplied by a single bronchiole. Most of the growth in the vascular system that occurs after birth includes further smooth muscle growth within the walls of arteries and arterioles and greater density and refinement of the arterioles and capillaries in the distal airway region.33,34 The thoracic wall in infants is more compliant, and their muscles are less developed than the muscles of adults and provide little structural support. The infant thoracic cage is also more boxlike, with the ribs being horizontally oriented or elevated (Figure 8-12). In addition, the diaphragm inserts into the thoracic cage in a horizontal plane, which decreases the effective ability to enlarge the thorax. Thoracic shape and dimension vary from individual to individual and are linked to age, gender, and race. At birth, the thorax has a smaller transverse dimension, which widens with the onset of walking. Thoracic size and volume continue to increase throughout childhood and especially during the adolescent growth spurt. However, development of the thorax and lung volume is not equal in both sexes. When evaluating lung size and volume throughout puberty and into adulthood, boys and men are consistently found to have larger lungs than age-matched and height-matched girls and women.40 Some races have a proportionately larger thorax-to-height ratio than others. In females, the location of the nipple varies with the size and shape of the breast. In males, the nipple is usually located in the midclavicular line at the level of the fourth intercostal space. Imaginary lines are commonly used to establish reference points and identify landmarks on the thorax. These lines and points help identify the location of underlying structures and the location of abnormal findings. On the anterior chest, the midsternal line divides the thorax into equal halves. The left and right midclavicular lines are parallel to the midsternal line. These are drawn through the midpoints of the left and right clavicles (Figure 8-13). The midaxillary line divides the lateral chest into equal halves. The anterior axillary line is parallel to the midaxillary line. It is situated along the anterolateral chest. The posterior axillary line is also parallel to the midaxillary line. It is located on the posterolateral chest wall (Figure 8-14). Three imaginary vertical lines are located on the posterior thorax. The midspinal line divides the posterior chest into two equal halves. The left and right midscapular lines are parallel to the midspinal line. They pass through the inferior angles of the scapulae in a relaxed upright subject (Figure 8-15). Anatomical Directions Descriptions of various anatomical structures often use the following terms: The thoracic cavity is formed by the tissues of the chest, upper back, and diaphragm.41 It is a cone-shaped cavity that houses the lungs and the contents of the mediastinum (Figure 8-16). It functions to protect the vital organs within and is capable of changing shape to enable air to be moved into and out of the lungs. The thoracic cavity is formed from epithelial, connective, and muscle tissues. The various parts of the thoracic wall are shown in Figure 8-17. The outer covering of the thorax is formed by the integumentary system, which includes skin, hair, subcutaneous fat, and breast tissues. Skin is a composite of an outer epidermis and an inner connective tissue layer called the dermis. Below the dermis is a layer of subcutaneous fat. Skeletal muscle, encased in a layer of connective tissue called fascia, is found under the subcutaneous fat. Skeletal muscle tissue forms the various muscles of the chest and back and lies over and between the ribs. The ribs of the rib cage lie in the inner portion of the thoracic wall. The inner layer of the thoracic wall is lined with a serous membrane called the parietal pleura. It is apposed by another serous membrane called the visceral pleura, which covers the lung. A thin, fluid-filled pleural space forms between the parietal and visceral pleural membranes. The rigidity of the thorax is provided by the bone tissue of the rib cage. The bony parts of the rib cage include the sternum, ribs, thoracic vertebral bones, scapula, and clavicle (Figure 8-18). The sternum is a long, vertical flat bone found on the anterior side that is composed of three bones: the manubrium, the body (or gladiolus), and the xiphoid process. The superior edge of the manubrium forms a shallow depression that is known as the suprasternal (or jugular) notch. The fused connection between the manubrium and the body is known as the sternal angle; it is also known as the angle of Louis. The sternal angle is an external marker of the point where the trachea divides into the left and right main stem bronchi. A cartilaginous joint called the costal cartilage is on the lateral edges of the manubrium and sternal body and forms the attachment between the ribs and sternum. This joint allows the rib cage to bend and permits the thorax to increase and decrease in size. The rib cage is formed by 12 pairs of ribs.41 Rib pairs 1 through 7 are known as the true ribs because they are attached directly to the sternum. The first ribs and the upper sternum form the opening into the thorax that is called the thoracic inlet, or operculum. Ribs 8 through 12 are called false ribs because they are either indirectly attached to the sternum or not attached at all. The vertebrochondral ribs include rib pairs 8, 9, and 10, which are indirectly attached to the sternum through a common cartilaginous strap. Rib pairs 11 and 12 are called floating ribs because they are not attached to the sternum. Each rib has a sternal end; a long, curved, and relatively flat body; and a head that articulates with the thoracic vertebrae (Figure 8-19). Intercostal muscles lie between the ribs and hold them together. Just below each rib is a thoracic artery, vein, and nerve that supply blood flow and nerve communications to that region of the chest wall (see Figure 8-17). The upper and lateral regions of the thorax house the bones of the pectoral girdles. The pectoral girdle on each side is formed by the clavicle and scapula.41 The scapula forms the socket for the shoulder joint and is stabilized or moved by skeletal muscles of the upper back. The clavicle supports and stabilizes the shoulder joint through a flexible attachment to the manubrium of the sternum. The various ribs move in different ways, and some may move more than others at different times. The first rib moves slightly, raising and lowering the sternum. Its slight motion increases the anteroposterior diameter of the chest. This action is not used during quiet breathing and becomes active only under conditions that require increased ventilation or deep breathing. Ribs 2 through 7 move simultaneously about two axes (Figure 8-20). As each rib rotates about the axis of its neck, its sternal end rises and falls. This movement increases the anteroposterior thoracic diameter in what is commonly referred to as a “pump handle”–like motion. At the same time, the rib moves about its long axis from its angle at the sternum. This motion causes the middle part of the rib to move up and down in what is commonly described as a “bucket handle.” The compound action of ribs 2 through 7 changes both the anteroposterior and the transverse dimensions in an upward and outward motion. Ribs 8 through 10 rotate in a pattern similar to that of ribs 2 through 7. However, elevation of the anterior ends of these ribs produces a small backward movement of the lower sternum that slightly reduces the thoracic anteroposterior diameter. Outward rotation of the middle section of these ribs increases the transverse diameter of the thorax. Ribs 11 and 12 participate in changing the contour of the chest in a minor way as they are pulled upward and outward in a “caliper”-like motion.
The Respiratory System
 State the major developmental events of the respiratory system.
State the major developmental events of the respiratory system.
 Describe how genes control lung development.
Describe how genes control lung development.
 Describe the key elements of normal fetal circulation.
Describe the key elements of normal fetal circulation.
 State what happens to the respiratory system at birth.
State what happens to the respiratory system at birth.
 Describe the developmental events in the respiratory system that continue after birth.
Describe the developmental events in the respiratory system that continue after birth.
 Identify the main structures in the thorax and describe their functions.
Identify the main structures in the thorax and describe their functions.
 Identify and describe the primary and accessory muscles of breathing.
Identify and describe the primary and accessory muscles of breathing.
 Describe how the pulmonary and bronchial circulations are organized and their functions.
Describe how the pulmonary and bronchial circulations are organized and their functions.
 Identify the major structures of the upper respiratory tract and how they function.
Identify the major structures of the upper respiratory tract and how they function.
 Describe how and why airways produce and move mucus.
Describe how and why airways produce and move mucus.
 Describe how the structures in the respiratory bronchioles and alveoli are organized.
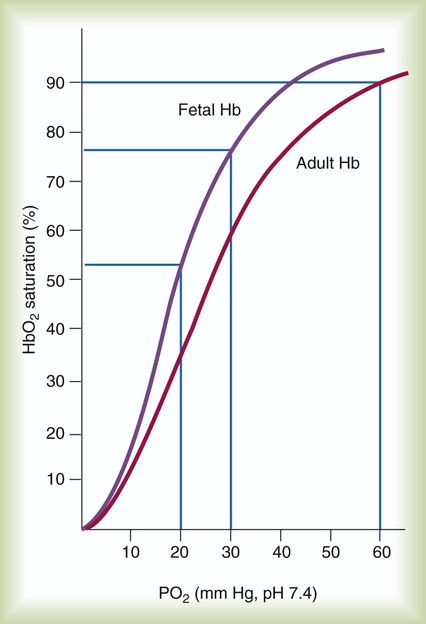
Describe how the structures in the respiratory bronchioles and alveoli are organized.
Development of the Respiratory System
Gestational Age
Developmental Event
Embryonic Period
20-22 days
Primordial pharyngeal arches form
21-23 days
Primordial respiratory cells form on fourth pharyngeal pouch, primordial heart starts forming
26th day
Laryngotracheal bud forms
4th wk
Primitive trachea develops
5th wk
Primary bronchial buds form, laryngeal structures develop
Fetal Period
Pseudoglandular Stage
6th wk
Segmental and subsegmental bronchioles form
7th wk
Diaphragm complete
8th wk
Heart complete, fetal circulatory pattern begins to develop
10th wk
Pulmonary lymphatic structures develop
12th wk
Major arteries formed
13th wk
Major airway epithelia and mucus-producing cells formed, smooth muscle cells developing
14th wk
Principal arteries formed
16th wk
Terminal bronchioles and associated pulmonary vessels form
Canalicular Stage
16th-17th wk
Respiratory bronchioles and immature acini begin to form
20th-24th wk
Type I and II pneumocytes begin to appear and replicate
24th-26th wk
Pulmonary capillaries develop at surface of acinus, immature surfactant begins to appear in lung fluid
Terminal Saccular Stage
26th wk-birth
Terminal saccules increase in number, pulmonary capillary density and proximity increase, type I and II pneumocytes continue to multiply, surfactant production increases, extrauterine life possible with support
Alveolar Stage
32th-40th wk
Immature alveoli begin to form and increase in number; surfactant production matures
40th week
50 million immature alveoli formed
Period After Birth
Birth
First breath and lung fluid cleared, adult circulatory pattern established
8-10 yr
470 million mature alveoli formed
Event
Factors and Genes
Early lung bud development and airway branching
Thyroid transcription factor 1 (NKX2-1)
FGF10 and FGF9
FGFR2IIIB and FGFR2IIIC
GATA-6
HNF-3 alpha and beta
Vitamin A (retinoic acid) and receptor
LEFTY 1 and 2
Sprouty (SPROUTY2)
Sonic hedgehog (SHH)
Bone morphogenetic protein 4 (BMP-4)
NOGGIN
N-Acetylglucosaminyltransferase 1
Secondary dichotomous branching
Transforming growth factor alpha and beta (TGF-α and TGF-β)
WNT 5 and 7
Platelet-derived growth factor (PDGF)
Alveolar development
PDGF
Tropoelastin 1
Fibrillin 1
Cyclin-dependent kinase inhibitors (p57 and p21)
Type 1 cell alpha transmembrane protein
Ephrin B2
Surfactant formation
Surfactant protein A, B, and C (SFTPA, SFTPB, and SFTPC)
ATP-binding cassette subfamily (ABCA3)
NKX2-1
HNF-3
Pulmonary vascular development
Activin receptor–like kinases
TGF
Vascular endothelial growth factors (VEGF)
Forkhead box transcription factors (Fox)
Integrin alpha forms
Caveolin 1 and 2
Transition from Uterine to Extrauterine Life
Placental Structure and Function
Value
Maternal Intervillous Blood
Fetal Umbilical Artery Blood
Fetal Umbilical Venous Blood
pH
7.38
7.36
7.39
PCO2 (mm Hg)
42
47
43
PO2 (mm Hg)
50
19
30
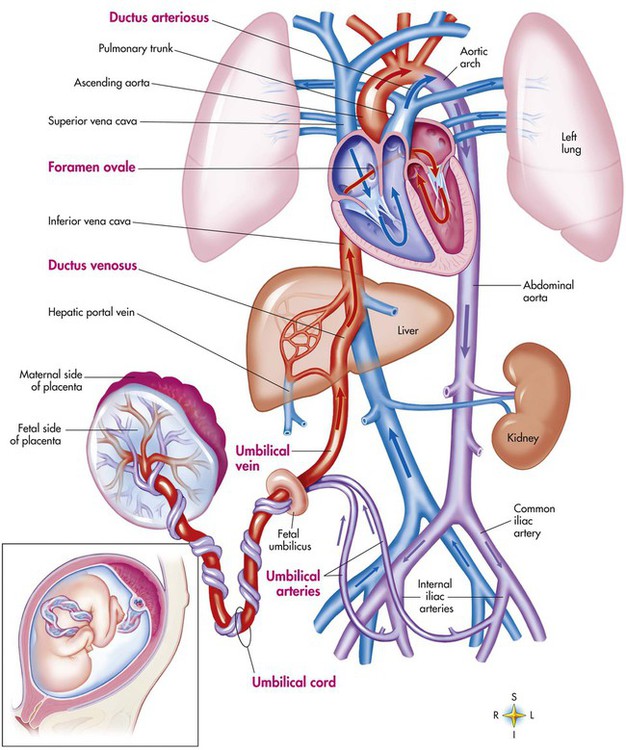
Fetal Circulation
Cardiopulmonary Events at Birth
Postnatal Lung Development
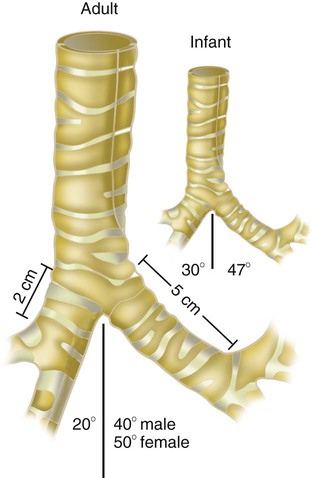
Upper Airway
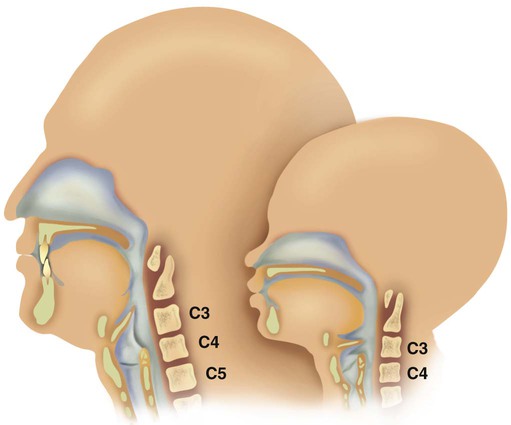
 to 3 months of age. Mechanical and chemical irritant laryngeal reflexes develop at birth and can initiate protective laryngeal closure; these reflexes can trigger prolonged apnea in some and may be a cause of sudden infant death syndrome.30 In addition, infections in this area or repeated attempts at intubation or suctioning can easily cause swelling and obstruction of this area.
to 3 months of age. Mechanical and chemical irritant laryngeal reflexes develop at birth and can initiate protective laryngeal closure; these reflexes can trigger prolonged apnea in some and may be a cause of sudden infant death syndrome.30 In addition, infections in this area or repeated attempts at intubation or suctioning can easily cause swelling and obstruction of this area.
Lower Airway and Alveoli
 postnatal years.33 This development largely occurs by the formation of increasing numbers of septa in the terminal airspaces that continue to subdivide the airspace into shallow immature alveoli. These immature alveoli enlarge in size and undergo further refinement of pulmonary capillaries over the ensuing months and years.34 By adulthood, the alveolar-capillary membrane has a gas exchange surface area of approximately 140 m2.35
postnatal years.33 This development largely occurs by the formation of increasing numbers of septa in the terminal airspaces that continue to subdivide the airspace into shallow immature alveoli. These immature alveoli enlarge in size and undergo further refinement of pulmonary capillaries over the ensuing months and years.34 By adulthood, the alveolar-capillary membrane has a gas exchange surface area of approximately 140 m2.35
Development of Vascular, Lymphatic, and Nervous Systems
Chest Wall Development, Diaphragm, and Lung Volume
Respiratory System in the Adult
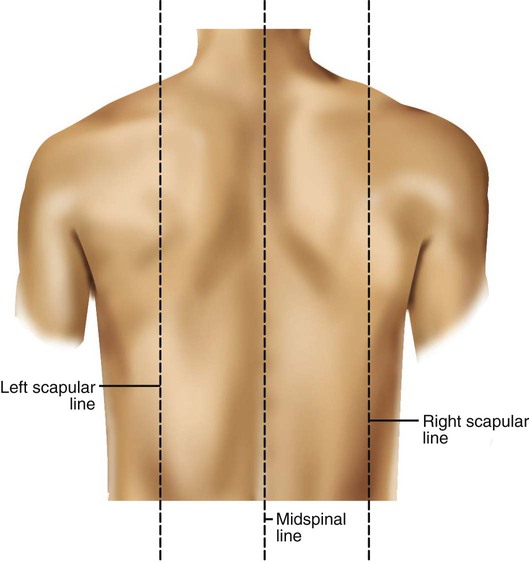
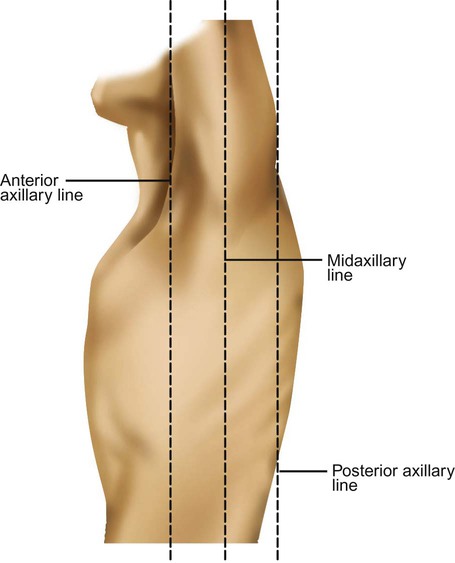
Surface Features of the Thorax
 Rule of Thumb
Rule of Thumb
Anterior, anteriorly
Front of the body, toward the front
Posterior, posteriorly
Back of the body, toward the back
Anteroposterior
In a direction from the front to the back
Lateral, laterally
Side of the body, toward the side
Medial, medially
Midline of the body, toward the midline
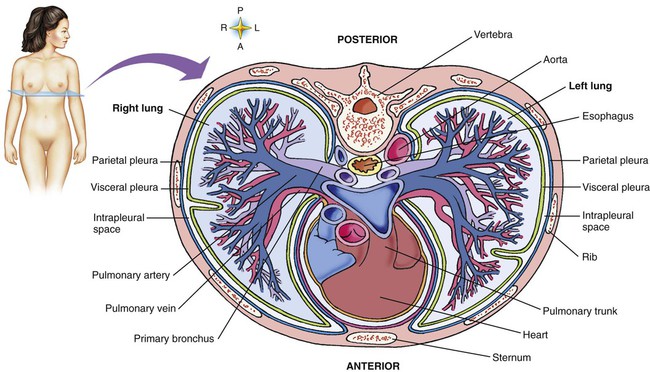
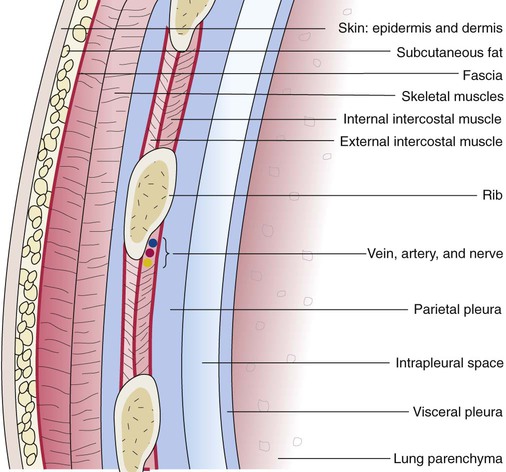
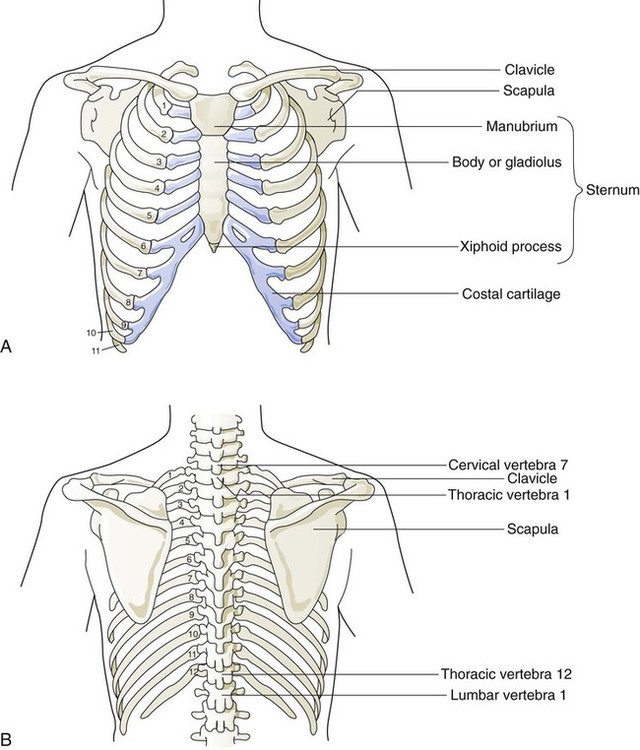
Components of the Thoracic Wall
Rib Movement
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


The Respiratory System



 Rule of Thumb
Rule of Thumb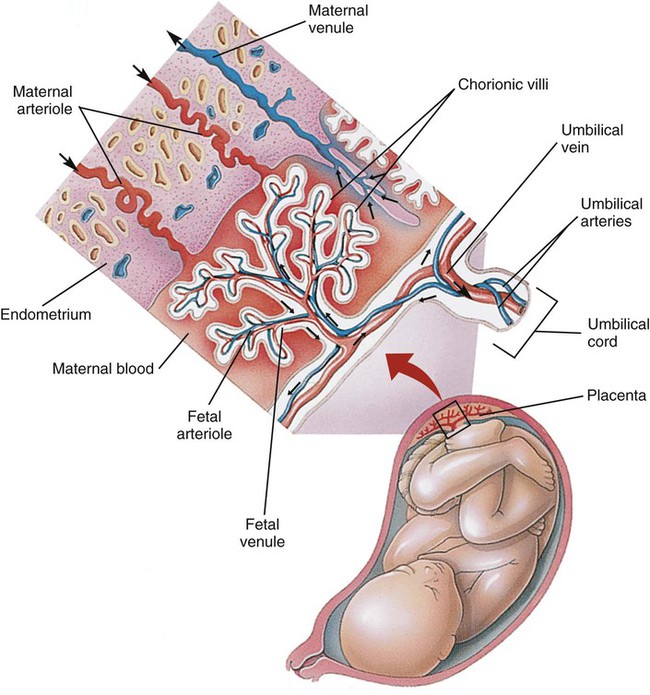
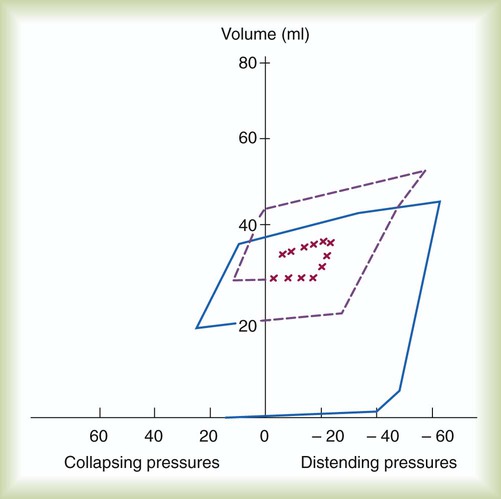
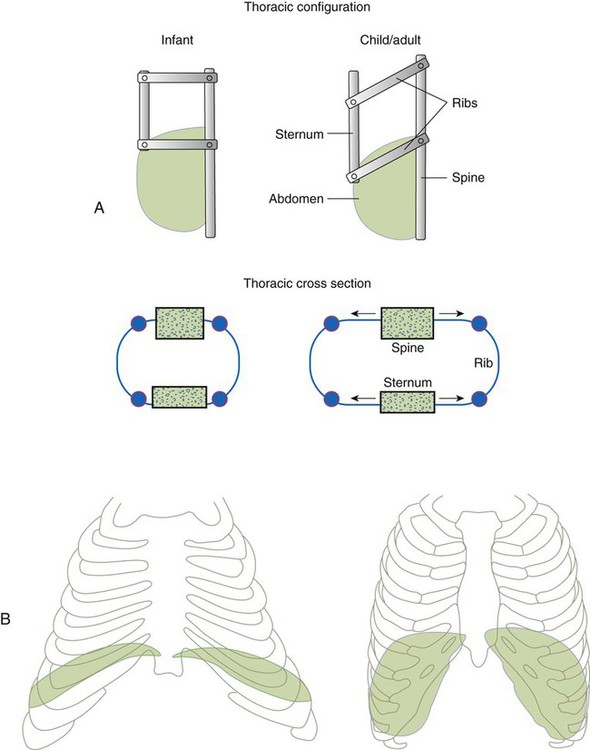
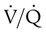 ) mismatch, and resultant hypoxemia. The combination of a reduced lung volume and high O2 consumption in an infant renders the infant more susceptible to profound hypoxemia in situations that disturb ventilation, lung volume, or
) mismatch, and resultant hypoxemia. The combination of a reduced lung volume and high O2 consumption in an infant renders the infant more susceptible to profound hypoxemia in situations that disturb ventilation, lung volume, or 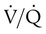 matching further. Infants possess a remarkable ability to elevate their lung volume dynamically. Infants, especially infants in distress, can actively increase lung volume by trapping gas, which improves
matching further. Infants possess a remarkable ability to elevate their lung volume dynamically. Infants, especially infants in distress, can actively increase lung volume by trapping gas, which improves 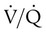 matching and gas exchange. Infants accomplish gas trapping actively by using the diaphragm during exhalation to slow expiration and to adduct (close) the vocal cords and narrow the glottis. The combination of these two maneuvers effectively regulates volume in the lung and dynamically elevates lung volume. The narrowing of the glottis or larynx during exhalation is referred to as “laryngeal braking.” Infants in respiratory distress commonly grunt, a manifestation of laryngeal braking. A more compliant chest wall contributes to suprasternal, substernal, intercostal, and subcostal retractions in distressed infants and young children (see Mini Clini).
matching and gas exchange. Infants accomplish gas trapping actively by using the diaphragm during exhalation to slow expiration and to adduct (close) the vocal cords and narrow the glottis. The combination of these two maneuvers effectively regulates volume in the lung and dynamically elevates lung volume. The narrowing of the glottis or larynx during exhalation is referred to as “laryngeal braking.” Infants in respiratory distress commonly grunt, a manifestation of laryngeal braking. A more compliant chest wall contributes to suprasternal, substernal, intercostal, and subcostal retractions in distressed infants and young children (see Mini Clini). Problem
Problem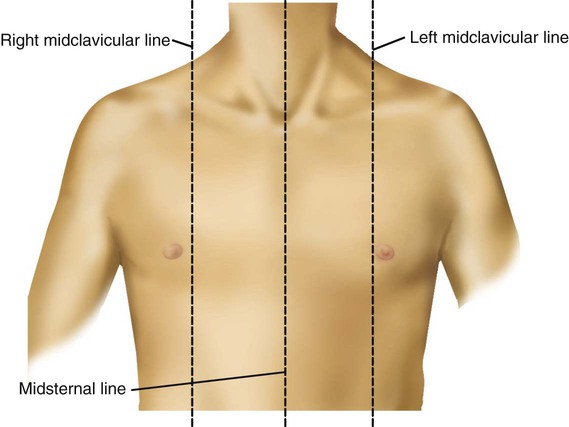
 Rule of Thumb
Rule of Thumb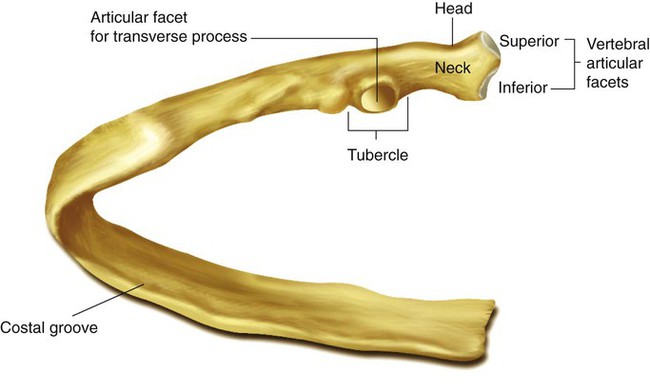
 Rule of Thumb
Rule of Thumb