7 The Biologic Pacemaker
In October 1958, the first fully internalized pacemaker was implanted at the Karolinska Institute in Sweden. Within hours, the unit ceased to function, or as Senning wrote in a retrospective account, “At 2 am the pacemaker became silent.”1 In a Medline search with the keywords “implantable pacemaker,” the first five citations (starting in 1960) are case reports of various implants, and the sixth (1962) is entitled “Complications of an implantable cardiac pacemaker.”2 Since that initial implant, millions of pacemakers have been implanted, saving lives and reducing or even eliminating symptoms. Along with these benefits of electronic pacemakers, however, complications occur in 5% to 10% of patients undergoing implant procedures.3,4 Acute complications range from minor hematomas to more severe infections, pneumothoraces, and cardiac perforations. Long-term management of pacemaker patients is complicated by device and lead failures and a requirement to change out the device when the battery fails. These limitations of device therapy have motivated the search for alternatives. One widely publicized approach over the last few years is to create a “biologic pacemaker” by either gene transfer or cell transplantation methods (Table 7-1). This chapter reviews the literature on this approach.
 Gene Therapy Approaches
Gene Therapy Approaches
Beta-2 Adrenergic Receptor Gene Transfer
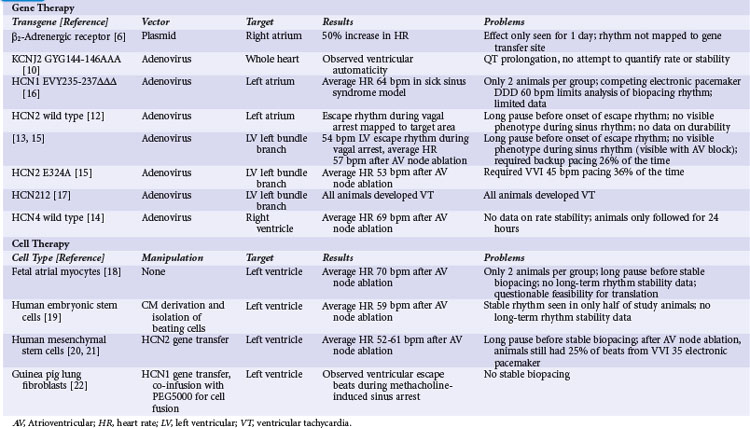
In the first attempt at genetic modulation of heart rate, Edelberg et al.5 injected plasmids containing the beta-2 adrenergic receptor (β2-AR) into mouse right atria. They found expression of the vector in 81% of myocytes at the injection site and a heart rate of 550 beats per minute (bpm) for the active-treatment group, significantly higher than the 370 bpm rate in controls. In a subsequent pig study, these investigators did not quantify gene transfer but reported a heart rate of 163 bpm for the β2-AR–receiving pigs relative to 127 bpm for controls.6
These studies showed promising results but also raised questions. The investigators did not report the time course of heart rate increase for the mice, but in the pigs they found a statistically significant increase in heart rate only on the second day after injection. The usual experience with plasmid-mediated gene transfer is that the effect persists for several weeks or even months after gene transfer.7–9 The transient effect for the β2-AR experiments suggests that some toxic effect from expression of this gene may be a limiting factor. Additional concerns involve possible mechanisms for the increase in heart rate. The mouse study did not target the specialized conducting system, and under ordinary circumstances, atrial myocytes should not display automaticity. A potential mechanism for the heart rate increase from β2-AR expression in atrial myocytes is Gs-mediated activation of adenylate cyclase, causing an intracellular cascade that ultimately results in phosphorylation of the type 1 calcium channel and the ryanodine receptor. Increased activity of these calcium-handling proteins could cause automaticity by the calcium clock mechanism, but sustained increase in intracellular calcium would likely be toxic. The porcine study did target sinus node by injecting in the area electrically mapped as the earliest site of activation during sinus rhythm, but the extremely transient functional change in pigs suggests that toxicity may be a concern even in sinus nodal tissues.
Ik1 Knockout
In the next illustration of heart rate modulation, investigators used a dominant negative mutation of the gene encoding the IK1 channel (KCNJ2 GYG144-146AAA) to create automaticity.10 IK1 (also called the inward rectifier current) is the principal current responsible for maintaining the cardiac myocyte’s resting membrane potential. The logic behind this strategy was that differences in IK1 between atrial and sinus nodal myocytes account for the relative instability of sinus nodal membrane potential, so that a deficit of IK1 would increase the probability of phase 4 depolarization and automaticity. The investigators admitted that their primary interest was in the electrophysiologic effects of IK1 on the action potential, and that they did not intentionally create pacemaking nodes. Their delivery method essentially scattered the transgene around the ventricular myocardium. Nonetheless, they found evidence of ventricular automaticity in 40% of animals expressing the KCNJ2-AAA mutation. Further investigation found that at least 80% of endogenous IK1 needed to be eliminated for automaticity to occur.11 There have been no subsequent reports applying this strategy to other models.
No attempt was made to quantify rate or stability of the idioventricular rhythm in this report.10 Other concerns rising from the KCNJ2-AAA strategy include the repolarization delaying effects of gene expression and the possibility of atrioventricular (AV) nodal suppressing effects. The investigators reported some level of action potential prolongation and increased QT interval at all expression levels of the transgene, as opposed to the pacing phenotype that was only visible with 80% suppression of IK1. Inspection of the limited reported data also shows no apparent AV nodal conduction, even in areas where the atrial beat is timed far enough away from the proceeding ventricular beat that AV nodal conduction should reasonably be expected. This finding is extrapolated from extremely limited data and should be assessed with more care if IK1 knockout becomes part of a biopacing strategy. Overall, the findings with IK1 knockout suggest that this may be a component of an overall biopacing strategy (with the previous caveats), but that the limited rate in pigs suggests that it might need to be applied with other interventions to increase automaticity.
If Manipulations
Since the early attempts at pacemaker induction, almost all subsequent gene transfer approaches have involved manipulation of If, the putative pacemaker (funny) current. The first attempts consisted of intramyocardial injections of adenovirus vectors encoding wild-type HCN2. The initial report targeted the left atrium,12 and a subsequent report showed gene transfer to the left bundle branch (LBB).13 In both reports, transgene function was not apparent during normal sinus rhythm. During vagal stimulation–induced sinus arrest, the transgene-expressing animals had a significantly higher escape rhythm than controls, and this rhythm mapped to the target area. Overall, the escape rate was inadequate to be a viable pacemaker replacement, and the transgene-driven rhythm could not be seen outside of vagal stimulation, so the investigators could not comment on stability of this rhythm over time.
An additional report with the HCN4 isoform suggested that automaticity was not limited to HCN2. Cai et al.14 injected adenoviruses encoding either wild-type HCN4 with green fluorescent protein (GFP) or GFP alone into porcine ventricles.14 They performed AV nodal ablation 3 to 4 days after gene transfer and measured the residual ventricular rate. The HCN4-expressing animals had a ventricular escape rate of 69 bpm, whereas the rate was only 41 bpm in the GFP controls. Pace mapping of the rhythm in the HCN4-expressing animals showed that the rhythm originated from the target region. The heart rate measurement was essentially an acute study, so no data on rate stability were provided.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


