Chapter 4 Pulmonary Circulation
Circulatory Structure
Pulmonary Circulation
The pulmonary arteries lie near and branch in unison with the airways in the bronchovascular bundle. They are much thinner than systemic arteries and have proportionately more elastic tissue in their walls. The walls of the arterioles, with a diameter less than 100 µm, are so thin relative to those of their systemic counterparts that fluid and gas can move across them. Within the gas-exchanging zone, the arterioles give rise to a network of pulmonary capillaries in the alveolar walls that is continuous throughout the lungs. They are so numerous that, when distended, blood flows almost as an unbroken sheet between the air spaces (Figure 4-1). “Sheet flow” reduces vascular resistance and optimizes gas exchange by creating a very large surface area, estimated at over 100 m2. When the transmural pressure difference between the inside and outside of the vessels is low, many of the capillary segments are closed, but flow switches among segments frequently as some open and others close. Nonflowing segments are rapidly recruited into the pulmonary vascular bed as needed to accommodate increased flow and may be further distended by an increase in transmural pressure. Both recruitment and distention of the pulmonary capillary bed reduce resistance to blood flow and help to maintain a low pressure in the face of increased blood flow. This low pressure allows the capillary-alveolar membrane to be very thin (approximately 1 µm), facilitating diffusion of respiratory gases between blood and alveoli. A red cell that follows a capillary path from the pulmonary artery to a vein may cross several alveoli, with the average transit time through the vessels engaged in gas exchange calculated to be approximately 0.75 second. The capillaries unite to form larger alveolar microvessels, which become venules and then veins that run between the lobules toward the hila, where upper and lower pulmonary veins from each lung empty into the left atrium.
Circulatory Physiology
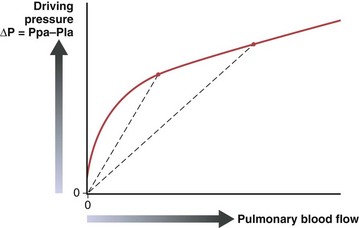
The pulmonary vascular resistance, PVR, is calculated as transvascular driving pressure, ΔP (mean upstream Ppa minus mean downstream Pla), divided by the flow: PVR = ΔP/Q. The calculated resistance must be interpreted in the context of flow, because the relationship of driving pressure to flow usually is not linear and its plotted curve does not pass through zero. As shown in Figure 4-2, pulmonary vascular resistance decreases as flow and pressure increase with the attendant recruitment and distention of vessels.
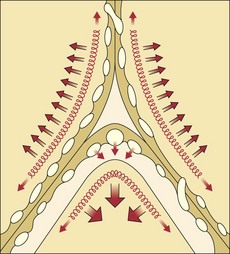
The resistance to flow through a vessel increases with its length, with the viscosity of the fluid, and, most important, with the inverse of the radius to the fourth power. In addition to muscle activity in the wall, the caliber of a distensible vessel depends passively on the transmural pressure difference between intravascular and extravascular pressures. This mechanism is particularly important in the lungs, where the vessels are embedded in expandable parenchyma. It is convenient to consider separately the effect of lung expansion on the extraalveolar arterial and venous vessels, which differs from the effect on the microvessels of the alveolar zone. With lung volume increase, extraalveolar vessels are distended as the pressure is lowered in the expanding perivascular space around them (Figure 4-3), and they are elongated as the lung expands.
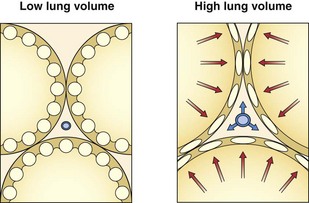
By contrast, the alveolar microvessels in the alveolar walls are elongated but partially collapsed by lung inflation, because the alveolar pressure that surrounds them tends to increase relative to the intravascular pressure. This effect is easy to recognize with positive-pressure ventilation, but it also occurs with spontaneous inspiration, because intravascular pressures fall relative to atmospheric and alveolar pressure. The sheets of capillaries in the alveolar walls are protected from the full compressive force of the alveolar pressure by the surface tension of the fluid that lines curved portions of the alveolar surface. Microvessels in the “corners” where alveolar walls meet are more fully protected from compression by the sharper curvature of the surface film and perhaps by local distending forces, analogous to the situation with extraalveolar vessels (Figure 4-4). The pulmonary vascular resistance is the sum of that through alveolar and extraalveolar vessels and thus has a complex relationship with lung volume. It is lowest at approximately the normal resting lung volume (functional residual capacity) but increases at higher and lower volumes.
Blood Flow Distribution
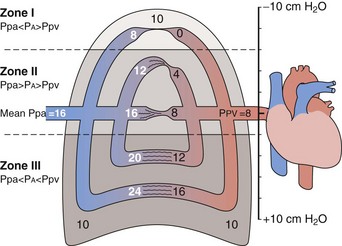
The gravitational effect has been conceptualized by dividing the lung into four zones, one above another, on the basis of the relationship of vascular and alveolar pressures (Figure 4-5). Intravascular pressures are higher at the bottom of the lung than at the top by an amount equal to a vertical hydrostatic column as high as the lung. Near the lung apex, zone I, the pressure in the alveoli (PA) exceeds that in both the pulmonary arteries (Ppa) and pulmonary vein (Ppv) and collapses the alveolar vessels, except those in the alveolar corners, which remain patent and allow some flow to continue. Below this, in zone II, Ppa exceeds PA, but PA is greater than Ppv, so flow depends on the pressure difference between Ppa and PA. The vessels remain open but are critically narrowed at the downstream end, where venous pressure is lower than alveolar pressure. This condition creates independence of flow from the downstream venous pressure, analogous to a waterfall in which a stream that flows over a precipice is unaffected by a rising level in the pool below until it rises above the level of the lip. In the middle to lower portion of the lung, zone III, both Ppa and Ppv exceed PA, the vessels are distended, and blood flow is the highest. Zone IV is restricted to a small area in the most dependent region, where flow diminishes. It has been postulated that this reduction is the result of increased vascular resistance secondary to low lung volume or perivascular edema in this area.
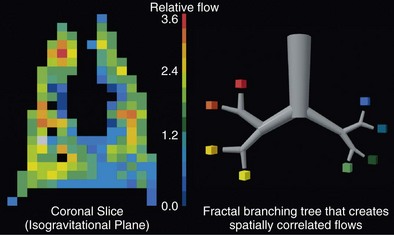
Although the gravitational effect expressed in the vertical zone concept contributes to the average increase in flow down the lung, it does not explain the observed large variability in flow within an isogravitational slice, which implies that other anatomic or vasoregulatory factors are important at this level. More recent studies have determined that the heterogeneous distribution of blood flow within horizontal (isogravitational) planes is due to asymmetric branching geometries (and hence resistances) of the vascular tree. Because the vascular tree is largely a dichotomous branching structure, differences in resistances between daughter branches cause flow to be distributed unevenly between the branches. With differences in resistances occurring at every bifurcation in the vascular tree, blood flow becomes progressively more heterogeneous, resulting in a broad distribution of flows at the terminal branches. Owing to the shared heritage up the vascular tree, neighboring lung regions have similar magnitudes of flow, with high-flow regions near other high-flow regions and low-flow regions neighboring other low-flow regions. Hence, the spatial distribution of pulmonary blood flow is not random but rather exhibits a clear pattern of high and low flows (Figure 4-6). Studies have demonstrated that the pattern of perfusion distribution is very stable over time and with growth, and that the pattern is genetically determined. These insights provide a new perspective on blood flow distribution in the lung. The traditional model of vertically stacked zones needs to be replaced by one in which the multiple zones can exist within horizontal planes. In addition, the large degree of heterogeneity within isogravitational planes suggests that mechanisms other than gravity must be responsible for the tight matching between regional ventilation and blood flow.