INTRODUCTION TO THE CONTROL OF RESPIRATION
Learning Objectives
The student will be able to summarize the neuroanatomical basis for the rhythm and pattern of simple respiration at rest.
The student will be able to describe the anatomy and physiology of both peripheral and central chemoreceptors.
The student will be able to distinguish the role that sensory afferent fibers sensitive to chemical/mechanical stimulation of the respiratory tract or muscle metabolism may have on ventilation.
The student will be able to define periodic breathing and discuss it in terms of abnormalities of either central or peripheral chemoreception.
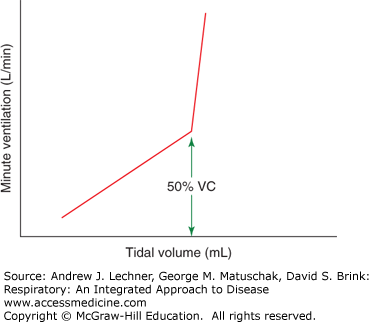
As demonstrated in earlier chapters, ventilation is simple in concept but complex in execution. The brain controls the basic pattern of breathing, integrating multiple influences within lower motor neurons of the brainstem and spinal cord to drive pharyngeal, laryngeal, diaphragmatic, intercostal, and other respiratory muscles. Recall that V̇E (L/min) = VT · f. The central nervous system regulates respiration by controlling the rhythm and pattern of its output to respiratory muscles, adjusting f, VT or both, depending on overall ventilatory needs for a greater or lesser V̇E (Fig. 11.1).
THE CENTRAL RHYTHM GENERATOR
The frequency of respiration, or its rhythm, is intrinsic to the brainstem. All vertebrates that use tidal oscillations for the exchange of O2 and CO2 in their lungs have such movements generated in their medulla oblongata. Indeed surgically isolated brainstem preparations, lacking afferent inputs from chemoreceptors and mechanoreceptors, still produce rhythmic outputs along the same cranial nerves as during normal respiration. One critical rhythm generator within a small area of the medulla and rostral to the obex is called the pre-Bötzinger complex. Bilateral lesions in this portion of the medulla induce complete respiratory arrest in humans. Whether this rhythmic discharge initiates within individual pacemaker cells, or from a network of such cells, remains a matter of debate. At this point, it is important to understand that a rhythm is constantly generated by the medulla, a rhythm that is modifiable by afferent input from sensory receptors.
THE CENTRAL PATTERN GENERATOR
The central pattern generator, or the brainstem output controlling all muscles involved in respiration, is much more complex. This pattern generator algebraically sums all the afferent inputs to produce well-coordinated activations of the diaphragm, intercostal muscles, and abdominal muscles, and if needed, the accessory muscles of respiration. Like respiratory rhythm, the goal of such pattern generation is maintenance of normal PaO2, Paco2, and pHa. Extreme examples of modulating both the rhythm and pattern of respiration are seen during rigorous exercise and ascents to high altitude. The pattern of respiration is also modulated by events like coughing, speech, sleep, vomiting, micturition, and defecation, particularly as the latter may mimic a Valsalva maneuver. Although some of these events are episodic or relatively infrequent, they can affect the normal respiratory pattern in dramatic ways.
DEFINED PATTERNS OF RESPIRATION
Eupnea: is normal, quiet breathing at rest. Individuals are usually unaware of it.
Tachypnea or polypnea: is an increase in f without an increase in VT. Tachypnea is not a normal stress response, unless hyperthermia or other factor has induced panting.
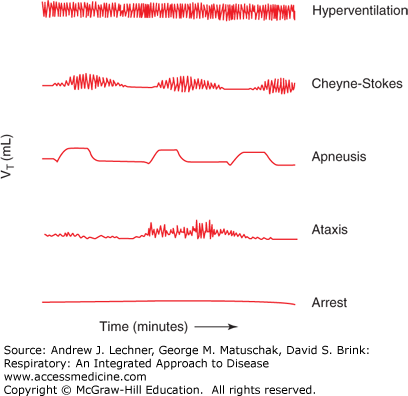
Hyperpnea or hyperventilation: denotes an increase in pulmonary ventilation involving both VT and f, but without the subjectively stressful sensations of dyspnea (Fig. 11.2).
Dyspnea: is the sensation of inadequate or stressful respiration, with exaggerated awareness of one’s need for increased respiratory effort. Dyspnea implies labored breathing, often involving accessory respiratory muscles. Many stimuli induce dyspnea.
Cheyne-Stokes respiration: is the most common form of abnormal breathing, with weak respiratory efforts that decrease to an apnea and then increase to hyperpnea. The “crescendo-decrescendo” oscillations of Cheyne-Stokes are most often caused by hypoxemia and are a frequently encountered symptom (see Chap. 25) (Fig. 11.2).
Apneusis: is an abnormally patterned breathing with prolonged inspirations that alternate with short expiratory movements (Fig. 11.2). Apneustic breathing is commonly noted after lesions in the pontine pneumotaxic center discussed below.
Ataxis or ataxic respiration: is an abnormal pattern with completely irregular breathing and increasing periods of apnea. As the pattern deteriorates, it may merge with agonal respiration. It is caused by damage to the medulla oblongata by stroke or trauma.
Apnea: is the absence of breathing. As generally used, apnea implies that the cessation is temporary. A prolonged apnea for any cause is considered respiratory arrest.
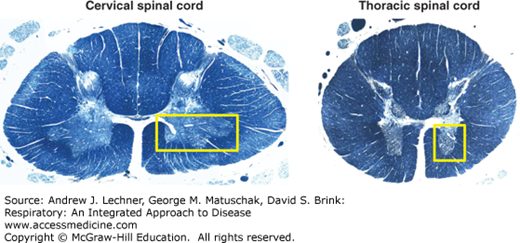
RESPIRATORY MOTOR NEURONS OF THE SPINAL CORD AND MEDULLA
Spinal cord neurons are organized into ventral horn (motor), dorsal horn (sensory), and lateral horn (autonomic) regions. Those in the ventral horn are most critical to respiration, since they include lower motor neurons innervating somatic striated muscles (Fig. 11.3). Major somatic striated muscles involved in respiration include:
Diaphragm: its innervating motoneurons exit at C3-C5 as the phrenic nerve.
Intercostal muscles: innervated by motoneurons within the thoracic ventral horn with axons that exit the spinal cord and distribute via the intercostal nerves.
Abdominal muscles: their motoneurons have axons that track within the lower thoracic and upper lumbar cord regions.
Accessory muscles: include all muscles that elevate and splay the ribs, notably the levator costalis, scalene, transverse thoracic, and sternocleidomastoid.
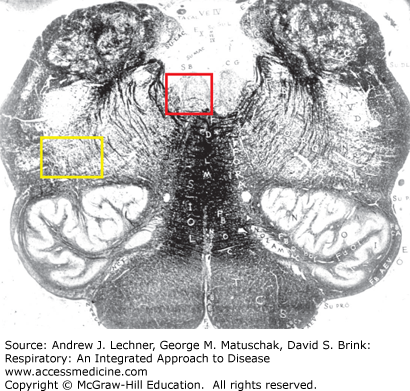
In addition to its role as the principal area of sensorimotor integration for respiration, the medulla contains lower motor neurons with fibers that exit via cranial nerves to innervate striated muscles in the head and neck (Fig 11.4). Motor neurons innervating the tongue muscles are found in the hypoglossal nucleus, while those innervating laryngeal, pharyngeal and facial muscles are found in the ventrolateral medulla. All of these muscles receive rhythmic central nervous system (CNS) motor input during every breath. Major cranial striated muscles that are involved during normal respiration include:
Laryngeal muscles: both laryngeal abductors and adductors are innervated by motoneurons in the nucleus ambiguus, whose axons travel with the vagus nerve.
Pharyngeal muscles: also receive motor input from the nucleus ambiguus by neurons whose axons exit via the glossopharyngeal and vagus nerves.
Facial muscles: most notably the m. nasalis, by motoneurons within the facial motor nucleus whose axons exit via the facial nerve.
Tongue muscles: principally the genioglossus muscle by motoneurons in the hypoglossal nucleus whose axons exit via the hypoglossal nerve.
Within the medulla are two recognized groups of neurons involved in the integration and coordination of breathing, whose functions continue to be intensively investigated. The first of these, the ventral respiratory column, is a longitudinal array of respiratory-related neurons that fire synchronously with each phrenic nerve discharge. These neurons are found in the ventrolateral reticular formation of the medulla, generally just ventral to the nucleus ambiguus. A subject’s basic respiratory rhythm persists if only these brainstem neurons are intact, albeit poorly controlled. Four main components of the ventral respiratory column have been identified (Fig. 11.5):
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


