INTRODUCTION TO THE NOMENCLATURE OF BIOLOGICAL CHANGE
Learning Objectives
The student will be able to describe the effects of cold, hot, and humid environments on the patterns of ventilation.
The student will be able to define cardiovascular drift and the experimental approach that is used to measure it.
The student will be able to enumerate changes in lung function after acute exposures to particulates, ozone, and other constituents of smog.
The student will be able to identify the changes in arterial blood gases, hyperventilation, and red cell mass with acute or chronic high altitude exposure.
The student will be able to explain the pulmonary physiology associated with breath-hold diving and deeper submersions using scuba gear.
Lung tissues and the processes of respiration are stressed when exposed to the extreme environments that humans inhabit. Although detailed discussions of each such environment are beyond the scope of this book, there are general respiratory responses that reflect the system’s adaptability over time, age, and level of aerobic capacity (Chap. 12). Situations that induce the most relevant adaptations include: cold, hot, and humid environments; regions with significant and persistent air pollution; short and long-term life at high altitude; and submersion under water using breath-holding or compressed air such as scuba diving gear. Each of these will be discussed here.
For clarity, this chapter recognizes three levels of organismal response to changing environmental conditions. Acclimation is a suite of responses induced in an organism by a single changed factor (like temperature, water supply, or light/dark cycle), usually in a laboratory setting where all other known variables are controlled. Periods of acclimation rarely last more than 2 months and involve individual organisms. For example, after spending a year living next to the airport you may barely hear its noise, but friends do not like to sleep over. Once you have moved away for a year, you will not sleep well at your old place either. Acclimations are transient and reversible.
Acclimatization is a more complex suite of responses by an organism to natural, multifactorial changes in the environment, perhaps encountered seasonally or when migrating across biomes. To achieve its full potential in the organism, acclimatization must optimize rapidly enough to yield benefits within one lifetime; fur density and lipid deposition only improve survival in the winter if they are in place by fall. Acclimatization usually involves at least a familial bloodline or other closely aligned group. Importantly too, larger groups undergoing acclimatization simultaneously have a higher likelihood of their changes enduring in the population. For example, families that bicycle or run regularly together often have similar body phenotypes. They are manipulating common pressure points within their similar genotypic backgrounds to affect more durable change within their own lifetimes. Any reader who owns animals in a temperate climate is familiar with their seasonal changes in girth, fur length, and breeding behaviors.
Knowing this, adaptation in a physiological sense is a durable response that has begun to integrate into the genome. Adaptation necessarily involves a population of organisms large enough to offer genotypic diversity to the selective pressures applied by an environment. In this sense of the word, adaptation is always at least partially genetic and inheritable. There are human groups that have lived at high altitudes (>3,500 m above sea level) in relative isolation for millennia. They share common physiques that differ from groups at sea level, such as greater chest circumference/height ratios, which their children show even if born and raised at sea level. Their erythropoietic responses to severe hypoxia differ from those of sea level natives. Indeed, hemoglobin diversity teaches some of the most interesting lessons in adaptive evolution for this reason.
CARDIOPULMONARY FUNCTION IN COLD ENVIRONMENTS
The severity of cold stress is determined by both ambient temperature and wind speed—what everyone appreciates as the “wind chill factor” in a weather report. Most heat is lost from the body while in the cold by conduction, that is, direct physical contact with colder substrates, and by convection, that is, heat transfer to colder substrates through the intervening air. Clearly, insulated clothing can limit most heat loss except from body surfaces like the face that are difficult to cover completely. Insufficient insulation while in the cold elevates V̇o2, a physiological response involving isometric contraction of voluntary muscles (shivering thermogenesis) and adrenergic stimulation of organs like brown adipose tissue (non-shivering thermogenesis). Thus V̇E must increase.
Interestingly, simply inhaling cold air to meet an increased V̇o2 has few effects on whole-body heat exchange, until metabolism significantly increases V̇E. Pharyngeal and upper airway temperatures decrease, particularly when breathing extremely cold air, through the mouth. However, temperatures in the lower respiratory tract remain normal. Thus most pulmonary functions are unaffected by breathing cold air during exercise, although asthmatics may experience bronchospasms when doing so (Chap. 21). Individuals who experience bronchospasm from cold air can show a >15% reduction in FEV1. The trigger for this hyperreactive airway response is unresolved. Older data implicated progressive water loss from the lower respiratory tract, but a 1999 study found that the amount of evaporative water loss did not correlate with severity of the obstructive response. Thus, dehydration of the lower respiratory tract does not appear to be the primary cause of cold-induced asthma.
If ambient cold reduces TB, aerobic capacity also declines versus at 37°C. Most of the decline reflects the dependence of tissue metabolism on TB, an effect more commonly observed in poikilothermic organisms with a labile TB. Lower tissue temperatures reduce the maximal force achieved by muscle contractions, thus restraining both Q̇ and power generated by voluntary muscles during exercise. Such hypothermia also increases HbO2 affinity (Chap. 3), causing a leftward shift of the HbO2 dissociation curve that promotes O2 uptake in the lung but decreases O2 release in the peripheral tissues. CaO2 is unaffected by cold stress or can increase slightly, while resting O2 delivery (= Q̇ · CaO2) increases to meet the needs of skeletal muscles that are shivering in the cold.
Such increases in resting Q̇ and muscle metabolism during cold stress may reduce the work capacity of remaining body elements, proportionally to the metabolic cost of maintaining thermal equilibrium in the cold. As example, envision a male runner wearing only a singlet and shorts while doing hill repeats when ambient conditions are –5°C and 30 km/h wind. Heat lost from his exposed body parts (head, upper torso, arms, hands) stimulates shivering thermogenesis in those muscles, although they do little to propel him up the hill. The demand for additional blood flow to these non-running muscles reduces the amount of Q̇ available to serve running muscles of the pelvis and legs. This runner is not likely to achieve his best times going up the hill because his leg power output is lower, although his Q̇max, aerobic capacity, and V̇Emax are essentially the same as the day he ran last month when ambient conditions were 15°C and 5 km/h wind.
Repeated exposure to breathing cold air during a season of exercising outdoors will result in only limited acclimation, because individuals rarely spend more than a few hours per day in harsh ambient conditions before retreating to milder thermal conditions indoors. There is no persuasive evidence that the lungs acclimatize or adapt to prolonged life in the cold versus the substantial changes that take place in the integument, skeletal muscles, and adipose tissues during chronic cold exposure. In animals like rodents that are continuously exposed to the cold, the non-pulmonary organ responses can seasonally increase V̇o2max, as by non-shivering thermogenesis due to increased brown fat deposition. As discussed in Chap. 12, the lungs of such acclimatized animals are still less likely than the heart to be the rate-limiting step of aerobic capacity.
CARDIOPULMONARY FUNCTION IN HOT AND HUMID ENVIRONMENTS
When resting in hot, humid environments [TA ≥30°C; relative humidity (RH) ≥70%] for less than an hour, lung function in healthy individuals is not appreciably altered versus spirometric volumes and FEV1 under more hospitable conditions (10°C ≤TA ≤25°C;RH ≤50%). As noted for cold exposure however, hot and humid environments do affect key components of the Fick equation (see Chap. 12). Sub-maximal exercise in the heat for more than 15 minutes causes significant water loss by sweating and by fluid shifts from plasma to tissues. Any rise in TB due to exercise or ambient thermal stress also redistributes blood flow away from working muscles to the skin for evaporative cooling. The progressive fall in plasma volume decreases ventricular preload and thus stroke volume (SV). This reduction in SV by thermal stress requires a compensatory rise in HR if Q̇max is to be maintained (see Fig. 13.1). This increase in HR to compensate for the decreased SV that ensues during long-term heavy exercise is termed cardiovascular drift.
FIGURE 13.1
Changes in metabolic performance during 45 minutes of cycling exercise when ambient temperature was 22°C (cool) or 36°C (heat). Data are means ± standard deviations. From LaFrenz AJ et al: Effect of ambient temperature on cardiovascular drift and maximal oxygen uptake, Med Sci Sports Exer Jun; 40(6):1065-1071, 2008.
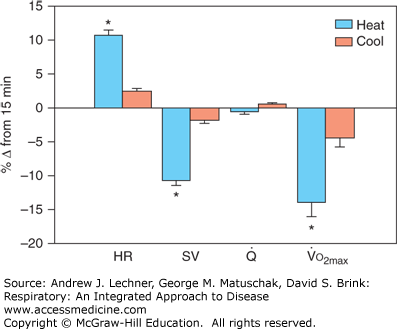
Not surprisingly, cardiovascular drift can be pronounced in the heat. Consider the runner again, now doing hill repeats when TA = 36°C and RH = 75% with no breeze. He cannot expect to achieve a personal best time today either, not because of lost aerobic capacity. Rather, there is again reduced work output by his running muscles, whose blood flow is being diverted to skin for evaporative cooling. Recent studies suggest a dramatic effect on cardiovascular drift and aerobic capacity of exercising during extreme elevations of TA. In a study by Lafrenz and colleagues (Fig. 13.1), the participants’ HR, SV, and Q̇ were measured after 45 minutes of strenuous cycling (60% of aerobic capacity) in either 22°C or in 35°C (RH = 40% in both). Despite adequate hydration in all athletes, TB increased by 1°C in the 35°C group versus 0.5°C in the 22°C group by 45 minutes, and showed cardiovascular drifts of 11% versus 2%, respectively. A stepwise exercise stress test (Chap. 12) conducted immediately after cycling found that aerobic capacity fell 15% in the 35°C group versus 5% in the 22°C cohort (Fig. 13.1).
Repetitive daily thermal stress induces heat acclimation, including an expansion of plasma volume by 5%-30% after prolonged intervals of heat exposure. When healthy persons were exposed to hot and dry conditions (TA = 50°C, RH ≤20%) for 2 hours per day for up to 2 weeks, their onset of sweating began at a lower TB versus their own pre-acclimated state, and their maximal sweat production (mg/m2/s) was threefold higher. Indeed their resting TB was 0.4°C lower after heat acclimation than before. As a result of this heat acclimation, their VT, f, and V̇E increased by 0.5 L/breath, 2 breaths/min, and 10 L/min, respectively, at an equivalent TB. These acclimation-induced changes in respiratory dynamics were associated with a PAo2 increase of about 4-5 mm Hg. Thus, repetitively performing exercise in the heat leads to an increase in V̇E commensurate with the enhanced rate of sweating when controlling for metabolic rate.
RESPIRATORY FUNCTION AND ENVIRONMENTAL POLLUTANTS
Periodic exposure to even low concentrations of ozone (0.2 ppm for 4 h/d × 2 d) can cause airway inflammation, leading to decreased FEV1 and FVC because of increased airway resistance. However, there is apparently natural variability in this responsiveness to ozone and to other constituents of smog. In more reactive individuals, ozone appears to directly activate airway irritant receptors (Chap. 11), leading to bronchoconstriction and contraction of distal smooth muscle within alveolar ducts. Such reactions decrease spirometric volumes and may or may not be reversible. Acclimatization to breathing in an environment with chronically poor air quality is conceivable. However, it has not been systematically observed in human populations or adequately modeled in animal studies.
Lung function is also affected by airborne debris, even ultrafine particles <0.5 μm in diameter (Chap. 10). Fine particulates are generated by machinery such as sand blasters and diesel engines, and can remain suspended for long periods of time. The amount deposited in upper and middle airways reflects the average V̇E during exposure, and thus increases with even brief periods of exercise. In healthy adults, abruptly breathing moderately clean air containing just 20,000 particles/cm3 can decrease FEV1 by 11 mL and FEF25-75 by 50 mL/s. The Environmental Protection Agency (EPA) reported that air collected within 20-50 m of major highways contained 300,000 particles/cm3. This was an amount 30-40 times higher than the EPA measured in parks or on lightly driven streets just a kilometer away. Thus, acute exposure to a large particulate burden can decrease a healthy person’s FEV1 by ~150 mL and FEF25-75 by 500 mL/s. In asthmatics, the risk of an adverse event from such exposure is greatly increased due to their hyper-reactive airways and enhanced susceptibility. Chronic exposure to air containing large particulate burdens causes lung injury similar to that noted with tobacco smoke and some occupational hazards. Whether human respiratory systems adapt over generations to poor air quality is an experiment in progress.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


