Chapter 6 Local regulation of vascular function
After reading this chapter, you should be able to:
Even in the absence of unusual structural design features like those discussed at the end of the last chapter, there is not always a straightforward relationship between regional microcirculatory blood flow and the applied pressure gradient. In many tissues, changes in perfusion pressure do not cause corresponding changes in flow because local adjustments of microcirculatory resistance tend to maintain blood flow constant (autoregulation). In addition, in some tissues with relatively low metabolic rates the blood flow is distributed unevenly across the capillary bed, with intermittent perfusion of capillaries in different areas (vasomotion). Finally, local vascular conductance is proportional to local tissue metabolism, with increased metabolic rate causing vasodilation and increased blood flow (functional hyperaemia). These three phenomena are due to a mixture of factors, the most universally important of which are the local vasodilator actions of chemicals released from the endothelium, local metabolites and the response of vascular smooth muscle cells to altered transmural pressure.
We will first examine the mechanisms involved in each of these three categories of local control.
ENDOTHELIAL CONTROL OF LOCAL CONDUCTANCE
Identity of endothelium-derived vasoactive factors
Endothelial cells release several factors that affect vascular smooth muscle cell activity. The most studied of these is the vasodilator nitric oxide (NO), endothelial synthesis of which is dependent on free intracellular Ca2+. NO acts by diffusing into the muscle cells and activating guanylate cyclase. There is good evidence that continual release of NO from the endothelium leads to a tonic dilator effect on the vascular smooth muscle, the magnitude of which depends on the balance between rate of release and rate of dispersal into the bloodstream. In addition, increased shear stress on the endothelium increases NO release. This probably constitutes an important factor in regulation of vessel calibre in large conduit arteries (see Chapter 7, p. 81) but, because of the large total cross-sectional area and concomitant low flow velocity in microvascular beds, the effect of shear stress on release of endothelial mediators in arterioles is likely to be much less substantial.
A number of additional biologically active factors of endothelial origin also exist, including EDRFs, that causes muscle cell hyperpolarization (endothelium-derived hyperpolarizing factor, EDHF) and for which there are several candidate molecules, together with the endothelins that cause vasoconstriction. None of these factors have been studied to the same extent as NO and their physiological significance is not yet clear, but it seems probable that at least collectively they may play a significant role in regulation of peripheral resistance (Quyyumi & Ozkor 2006).
Assessment of endothelial control of local conductance
Reactive hyperaemia
If the arterial inflow to a tissue is occluded for more than a few seconds, the conduit and microcirculatory arterial vessels dilate downstream of the occlusion. As a result, release of the occlusion is accompanied by a temporary elevation of local blood flow above the pre-occlusion level, termed reactive hyperaemia. The reactive hyperaemic responses of both conduit and microcirculatory vessels are attenuated after administration of substances that prevent NO production, suggesting that both involve EDRFs and might, therefore, be utilized as indices of endothelial dilator function. It is uncertain whether EDRFs other than NO are implicated.
Reactive hyperaemia is usually elicited in the forearm with occlusion of the brachial artery above the elbow for a period of between 2 and 3 min. In these circumstances, the peak flow immediately after release of occlusion is typically two to four times the pre-occlusion value and returns to this resting level over 30–50 s. Conduit vessel dilation is assessed using ultrasound imaging of brachial artery diameter, and the reactive hyperaemic response is referred to as flow-mediated dilation. Microcirculatory dilation is usually assessed either by measurement of blood flow through forearm or finger using venous occlusion plethysmography (Fig. 6.1) or by laser-Doppler measurement of forearm skin flow.
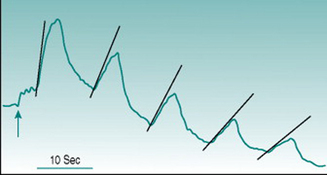
Comparative studies show poor correlation between the magnitudes of large and small vessel responses in different subject populations (Hansell et al 2004), which is perhaps not surprising if we consider the mechanisms likely to underlie dilation in the two vessel types. Assuming that all endothelial cells release EDRFs continuously, local accumulation of these factors during the period of arterial occlusion should have similar dilator consequences in large and small vessels once the normal pressure gradient is re-established. In conduit vessels, however, the increased volume flow will be accompanied by a high rate of shear stress on the endothelium that will increase local EDRF release further. As flow velocity is much lower in the microcirculation, shear stress is not likely to generate factor release at that site. A second complicating factor arises from the fact that cessation of blood flow through a tissue results in local accumulation of dilator metabolites (see Chapter 6, p. 67). Depending on the metabolic rate of the tissue and the duration of arterial occlusion, metabolic dilation in the microcirculation may, therefore, act additively with EDRFs.
Endothelial turnover
The endothelium is turning over continually, with dying cells being shed into the circulation and being replaced by new cells that originate from circulating bone marrow-derived progenitors. These processes may provide additional indices by which the functional status of the endothelium can be assessed at a whole-body level. On the one hand, large numbers of dislodged endothelial cells in the blood stream could indicate reduced viability, consistent with reduced EDRF function. A number of studies have reported elevated numbers of circulating endothelial cells (CECs) in various disease states associated with other evidence of reduced endothelial dilator capacity, including hypertension and diabetes. Conversely, it could be argued that rapid cell turnover would lower the average age of the endothelial population and so reduce the proportion of old, metabolically sub-optimal cells. This possibility is consistent with observations that CECs rise in parallel with endothelial dilator capacity in healthy subjects during aerobic training (see Chapter 11, p. 134).
CEC numbers alone, therefore, provide only ambiguous information on endothelial status and additional markers of endothelial viability must be used concomitantly in order to decide whether high CEC values denote healthy or unhealthy endothelium. One marker that may be useful is Von Willebrand factor, which is released from endothelial cells in response to inflammatory stimuli. Von Willebrand factor levels appear to be elevated in at least most pathological situations where CEC numbers are increased. By contrast, the one study that has compared these parameters with fitness levels in healthy subjects (O’Sullivan 2003) found that the rise in CECs associated with training was not linked to any change in plasma Von Willebrand levels.
Assessment of nitric-oxide involvement in endothelial control of local conductance
Release of nitric oxide
It can be anticipated that NO released from endothelium will diffuse into the bloodstream as well as into the vascular wall, so that plasma levels of NO could provide an index of endothelial dilator function. Detection of NO itself is not practicable, since the NO molecule is metabolized too rapidly to allow blood sampling and assay. However, the main breakdown product of NO, nitrite ion, is stable and recent studies have demonstrated good correlation between serum nitrite levels and other markers of NO-dependent dilation (Rassaf et al 2006).



