Chapter 9 Diffusion of respiratory gases
 For gas to transfer between the alveolus and the haemoglobin in the red blood cell it must diffuse across the alveolar and capillary walls, through the plasma and across the red cell membrane.
For gas to transfer between the alveolus and the haemoglobin in the red blood cell it must diffuse across the alveolar and capillary walls, through the plasma and across the red cell membrane.


Nomenclature in this field is confusing. In Europe, measurement of the passage of gases between the alveoli and pulmonary capillaries is referred to as lung ‘transfer factor’ (e.g. Tlco represents lung transfer factor for carbon monoxide). However, the older term ‘diffusing capacity’ (e.g. Dlco for lung diffusing capacity for carbon monoxide) which has been used for many years in the USA is now the recommended term1 despite the fact that some of the barrier to oxygen transfer is unrelated to diffusion (see below).
Fundamentals of the Diffusion Process
Diffusion of a gas is a process by which a net transfer of molecules takes place from a zone in which the gas exerts a high partial pressure to a zone in which it exerts a lower partial pressure. The mechanism of transfer is the random movement of molecules and the term excludes both active biological transport and transfer by mass movement of gas in response to a total pressure difference (i.e. gas flow as occurs during tidal ventilation). The partial pressure (or tension) of a gas in a gas mixture is the pressure which it would exert if it occupied the space alone (equal to total pressure multiplied by fractional concentration). Gas molecules pass in each direction but at a rate proportional to the partial pressure of the gas in the zone from which they are leaving. The net transfer of the gas is the difference in the number of molecules passing in each direction, and is thus proportional to the difference in partial pressure between the two zones. Typical examples of diffusion are shown in Figure 9.1.
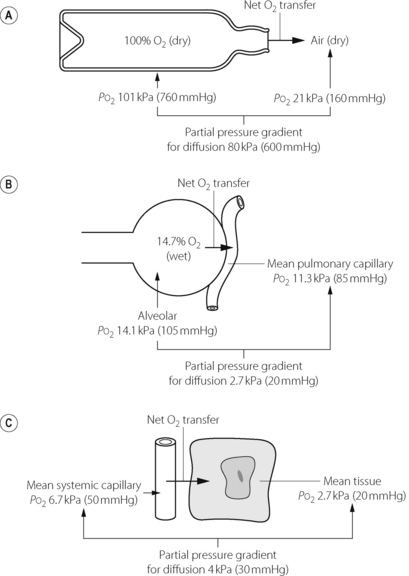
In each of the examples shown in Figure 9.1, there is a finite resistance to the transfer of the gas molecules. In Figure 9.1A, the resistance is concentrated at the restriction in the neck of the bottle. Clearly, the narrower the neck, the slower will be the process of equilibration with the outside air. In Figure 9.1B, the site of the resistance to diffusion is less circumscribed but includes gas diffusion within the alveolus, the alveolar/capillary membrane, the diffusion path through the plasma, and the delay in combination of oxygen with the reduced haemoglobin in the red blood cell (RBC). In Figure 9.1C, the resistance commences with the delay in the release of oxygen by haemoglobin, and includes all the interfaces between the RBC membrane and the site of oxygen consumption in the mitochondria. There may then be an additional component in the rate at which oxygen enters into chemical reactions.
In the living body oxygen is constantly being consumed, while carbon dioxide is being produced, so equilibrium cannot be attained as in the case of the open bottle of oxygen in Figure 9.1A. Instead, a dynamic equilibrium is attained with diffusion down a gradient between the alveolus and the mitochondria for oxygen and the reverse for carbon dioxide. The maintenance of these partial pressure gradients is, in fact, a characteristic of life.
Quantification of Resistance to Diffusion
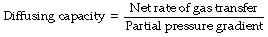
The usual biological unit of diffusing capacity is ml.min−1.mmHg−1 or, in SI units, ml.min−1.kPa−1.
Diffusion of Oxygen in the Lungs
It is now widely accepted that oxygen passes from the alveoli into the pulmonary capillary blood by a passive process of diffusion according to physical laws, though for a while it was believed that oxygen was actively secreted into the blood (page 242). It is believed that diffusion equilibrium is very nearly achieved for oxygen during the normal pulmonary capillary transit time in the resting subject. Therefore, in these circumstances, the uptake of oxygen is limited by pulmonary blood flow and not by diffusing capacity. However, when exercising, while breathing gas mixtures deficient in oxygen or at reduced barometric pressure, the diffusing capacity becomes important and may limit oxygen uptake.
Components of the Alveolar/Capillary Diffusion Pathway
The gas space within the alveolus. At functional residual capacity, the diameter of the average human alveolus is of the order of 200 μm (page 20), and it is likely that mixing of normal alveolar gas is almost instantaneous over the small distance from the centre to the periphery. Precise calculations are impossible on account of the complex geometry of the alveolus, but the overall efficiency of gas exchange within the lungs suggests that mixing must be complete within less than 10 ms. Therefore, in practice it is usual to consider alveolar gas of normal composition as uniformly mixed.
This generalisation does not seem to hold when subjects inhale gases of widely different molecular weights. This was first demonstrated in normal subjects inhaling mixtures of sulphur hexafluoride (SF6) and helium when the SF6 concentration was found to be higher (relative to helium) earlier in the breath.2 According to Graham’s law SF6 (molecular weight 146), would diffuse six times less readily than helium (molecular weight 4) and would therefore tend to remain concentrated at the core of the alveolus. A similar problem is seen with inhaled anaesthetic agents, for example a large proportion of the end expiratory/arterial partial pressure gradient for the anaesthetic isoflurane (molecular weight 184.5) cannot be explained by alveolar dead space or shunt and may be due to failure to achieve uniformity within the alveolus.3 Nevertheless, it seems unlikely that non-uniformity within a single alveolus is an important factor limiting diffusing capacity under normal conditions with gases such as oxygen, nitrogen and carbon dioxide, which have molecular weights that are not greatly different.
Alveolar lining fluid. Alveoli contain a thin layer of surfactant rich fluid (page 218) through which respiratory gases must diffuse.4 The depth of this fluid layer, and therefore its impediment to diffusion, is very variable. There are ‘pools’ of fluid in alveolar corners (see Figure 2.8) and in the depressions between where the capillaries bulge into the alveolus, with only a very thin layer on the surface of the capillary bulges, thus providing the minimal diffusion barrier in the most vital area.
Tissue barrier. Electron microscopy reveals details of the actual path between alveolar gas and pulmonary capillary blood, shown in Figure 2.7. Each alveolus is lined with epithelium which, with its basement membrane, is about 0.2 μm thick, except where epithelial cell nuclei bulge into the alveolar lumen. Beyond the basement membrane is the interstitial space, which is very thin where it overlies the capillaries, particularly on the active side; elsewhere it is thicker and contains collagen and elastic fibres. The pulmonary capillaries are lined with endothelium, also with its own basement membrane, which is approximately the same thickness as the alveolar epithelium, except where it is expanded to enclose the endothelial cell nuclei. The total thickness of the active part of the tissue barrier is thus about 0.5 μm, containing two pairs of lipid bilayers separated by the interstitial space.
Plasma layer. Human pulmonary capillaries are estimated to have a mean diameter of 7 μm, similar to the diameter of a RBC, part of which is therefore forced into contact with the endothelial cell surface (see Figure 2.7). The diffusion path through plasma may therefore be very short indeed, but only a small proportion of the RBC surface will be in such close proximity with the endothelium, much of the RBC passing through the middle of the capillary, up to 3.5 μm from the endothelial cell. Furthermore, since the diameter of the capillary is about 14 times the thickness of the tissue barrier, it is clear that the diffusion path within the capillary is likely to be much longer than the path through the alveolar/capillary membrane. A complex pattern of diffusion gradients is therefore established within the plasma depending on the oxygen tension in the alveolus and the number of RBCs present.5 This is discussed in more detail below with respect to carbon monoxide.
Diffusion into and within the RBC.6 Confining haemoglobin within the RBC reduces the oxygen diffusing capacity by 40% in comparison with free haemoglobin solution.7 There are three possible explanations for this observation. First, there is evidence that the rapid uptake of O2 and CO by RBCs causes depletion of gas in the plasma layer immediately surrounding the RBC.8 Referred to as the ‘unstirred layer’, this phenomenon is most likely to occur at low packed cell volume (PCV) when adjacent RBCs in the pulmonary capillary have more plasma between them.9 Secondly, oxygen must diffuse across the RBC membrane, though this is not normally believed to be a significant diffusion barrier. Thirdly, once in the cell, oxygen must diffuse through a varying amount of intracellular fluid before combining with haemoglobin, a process that is aided by mass movement of the haemoglobin molecules caused by the deformation of the RBC as it passes through the capillary bed, in effect ‘mixing’ the oxygen with the haemoglobin.
RBCs change shape as they pass through capillaries (both pulmonary and systemic) and this plays an important role in the uptake and release of oxygen.6 The dependence of diffusing capacity on RBC shape changes may result from reducing the unstirred layer by ‘mixing’ the plasma around the RBC, from changes in the cell membrane surface area to RBC volume ratio or from assisting the mass movement of haemoglobin within the cell. This has led to further studies in which the deformability of RBCs is reduced (using chlorpromazine) or increased (using sodium salicylate), which have demonstrated that diffusing capacity is increased with greater RBC deformability.9 Of more clinical significance is the effect of plasma cholesterol on RBC function.10 Elevated cholesterol concentration in the plasma causes increased cholesterol in the RBC membrane, a change that is known to make the membrane thicker and less deformable, both of which lead to reduced efficiency of diffusion across the membrane. Oxygen uptake by RBCs in the lung, and its release in the tissues, are both believed to be significantly impaired by hypercholesterolaemia, particularly in tissues with high oxygen extraction ratios such as the heart.
Uptake of oxygen by haemoglobin. The greater part of the oxygen that is taken up in the lungs enters into chemical combination with haemoglobin. This chemical reaction takes a finite time and forms an appreciable part of the total resistance to the transfer of oxygen.11 This important discovery resulted in an extensive reappraisal of the whole concept of diffusing capacity. In particular, it became clear that measurements of ‘diffusing capacity’ did not necessarily give an indication of the degree of permeability of the alveolar/capillary membrane.
Quantification of the Diffusing Capacity for Oxygen
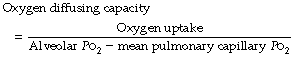
The alveolar Po2 can be derived with some degree of accuracy (page 140) but there are very serious problems in estimating the mean pulmonary capillary Po2.
The earliest analysis of the problem was made by Bohr in 1909.12 He made the assumption that, at any point along the pulmonary capillary, the rate of diffusion of oxygen was proportional to the Po2 difference between the alveolar gas and the pulmonary capillary blood at that point. Using this approach, and assuming a value for the alveolar/pulmonary end-capillary Po2 gradient, it seemed possible to construct a graph of capillary Po2, plotted against the time the blood had been in the pulmonary capillary. A typical curve drawn on this basis is shown as the broken line in Figure 9.2A. Once the curve has been drawn, it is relatively easy to derive the mean pulmonary capillary Po2, which then permits calculation of the oxygen diffusing capacity. The validity of the assumption of the alveolar/pulmonary end-capillary Po2 gradient is considered below.
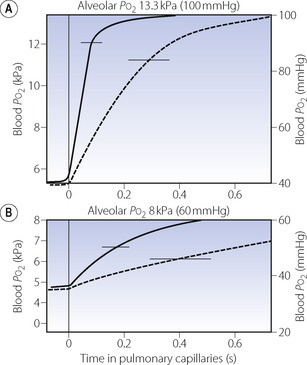
Fig. 9.2 Each graph shows the rise in blood Po2 as blood passes along the pulmonary capillaries. The horizontal line at the top of the graph indicates the alveolar Po2 that the blood Po2 is approaching. In (A) the subject is breathing air, while in (B) the subject is breathing about 14% oxygen. The broken curve shows the rise in Po2 calculated according to the Bohr procedure on an assumed value for the alveolar/end-capillary Po2 gradient. The continuous curve shows the values obtained by forward integration.13 Horizontal bars indicate mean pulmonary capillary Po2 calculated from each curve.
 Hb4(O2)4) has a much higher velocity constant than that of the combination of the other three molecules. This is discussed further on page 190.
Hb4(O2)4) has a much higher velocity constant than that of the combination of the other three molecules. This is discussed further on page 190.Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


