Chapter 9 Circulatory limits to acute exercise
After reading this chapter, you should:
Dynamic work capacity is proportional to the amount of aerobic metabolism that can take place in the cells of the locomotor skeletal muscles. Although the efficiency of these metabolic processes can be manipulated by, for example, dietary selection so as to optimize the appropriate substrates and training-induced adaptation of muscle fibre type, the end result is a reflection of oxygen availability. As such, the primary limit to exercise is the rate at which oxygen can be delivered to the muscle.
As discussed in Chapter 8 (p. 99), ventilatory function and pulmonary oxygen uptake in the pulmonary capillaries are not normally limiting factors since, even at peak workloads, arterial oxygen saturation is similar to that at rest, except in the subset of elite athletes with exceptional cardiac adaptations to training. However, we saw in Chapter 3 that the volume of blood that can be pumped around the circulation per unit time is determined by the age-limited maximum heart rate and by the decline in ventricular filling at high heart rates. These changes set an absolute limit of around 0.3 l/min/kg to cardiac output in an untrained individual, equating to around 21 l/min in a 70 kg person. The purpose of the current chapter is to examine in detail the factors that limit cardiac output at different exercise durations and intensities, under different environmental circumstances and in relation to different types of individual.
FACTORS THAT LIMIT MUSCLE PERFUSION
Effects of exercise hyperaemia
In resting muscle, where there is a relatively high precapillary vasoconstrictor tone, mean capillary hydrostatic pressure and plasma oncotic pressure are both around 25 mmHg, so that there is a balance between inwardly and outwardly directed pressure gradients and no net water movement occurs across the capillary wall. When the muscle arterioles dilate with onset of exercise, however, the resulting fall in precapillary resistance automatically raises capillary hydrostatic pressure, to values as high as 50–60 mmHg when the arterioles are maximally dilated. Because this is far above the opposing oncotic pressure value, net extravasation of plasma water takes place until the resultant increase in interstitial hydrostatic pressure is sufficient to rebalance the outward/inward forces (Fortney et al 1981). At least at high muscle workloads, the outward water movement is increased further by the rise in interstitial osmolality caused by accumulation of muscle metabolites.
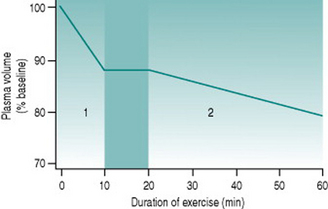
Since skeletal muscle makes up such a large proportion of total body mass, the volume of extravasated fluid within the muscles during whole-body dynamic exercise can be appreciable. At relatively heavy workloads, 10–12% of plasma volume (around 300 ml in a 70 kg person) is lost over the initial 10 min of activity (Fig. 9.1). Thus, over this period cardiac output also falls by around 300 ml provided heart rate remains constant, necessarily reducing muscle blood flow. Moreover, plasma loss results in a concomitant rise in haematocrit, elevating blood viscosity and so increasing cardiac workload.
Effect of heat production
As mentioned in Chapter 6 (p. 68), the first of these processes is withdrawal of sympathetic vasoconstrictor tone from the cutaneous arterioles. In parallel, there is activation of cholinergic sympathetic nerves to eccrine sweat glands. Both the loss of sympathetic tone and the process of sweat production cause cutaneous vasodilation. As a result, skin blood flow rises from a resting value that is typically less than 500 ml/min at environmental temperatures below 25° C to as much as 3 l/min after 15–20 min of sustained heavy exercise, with a similar fall in muscle blood flow. The dilatation due to sweating is at least partly secondary to glandular release of kinins (see Chapter 6, p. 75) but may also involve release of unknown dilator factors from the sudomotor nerve endings (Joyner & Halliwill 2000). As well as serving to convey heat to the body/environment interface, the high skin blood flow is essential to provide water and electrolytes for sweat production. This typically reaches a maximum of around 1 l/h, but may be as high as 2 l/h in a person who exercises routinely in hot conditions.
Sweating produces a second phase of plasma volume reduction that typically becomes measurable around 20 min after commencement of exercise (Fig. 9.1). Although in theory plasma and interstitial fluids are in equilibrium, high rates of sweat production draw primarily on the plasma, so that blood volume falls and haematocrit rises progressively as the exercise bout continues. Both effects make muscle oxygenation progressively more and more inefficient.
Measurement of sweating
A wide range of techniques are available for measuring sweat production, but none are suitable for all applications. Choice of technique depends critically on the intensity of sweat secretion to be measured and the duration of measurement required. All techniques in routine use involve one of four basic principles, as summarized below.
Volume collection
Except in very humid environments, sweat evaporates rapidly from the skin, even at quite high rates of secretion. If, however, a container is placed on the skin surface so as to prevent evaporative loss, then the sweat produced over that area of skin will be retained and can be measured. For obvious reasons, this technique cannot be used to detect the onset of sweating and is not practicable with very low sweat rates. But at high rates of production it can provide accurate quantitative values, provided that evaporative loss is prevented during the process of measurement and that the duration of collection is appropriate for the volume of the sampling vessel. One advantage of this method is the capacity to make repeated measurements for the same set of sweat glands over a prolonged period. A limitation is the lack of information obtained on behaviour of sweat glands elsewhere on the body.



