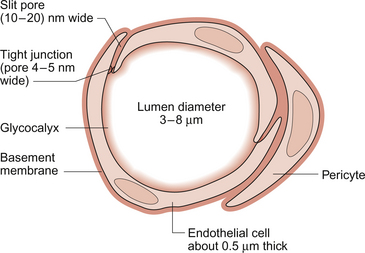
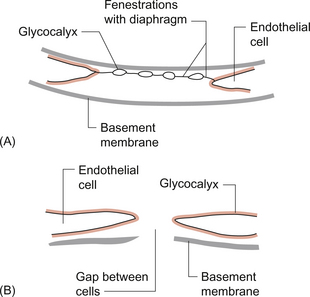
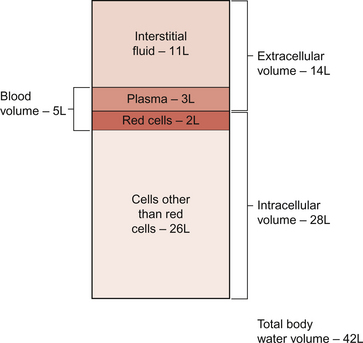
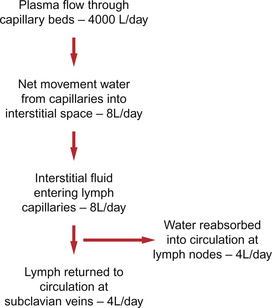
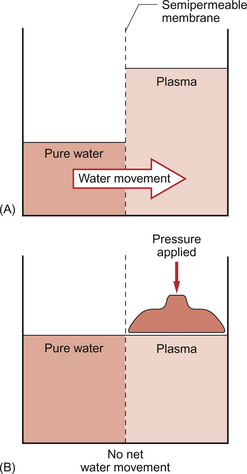
11 Aspects of the structure of capillaries are discussed, along with the other types of blood vessel, in Chapter 1. A diagram of a microcirculatory unit is shown in Figure 1.9. A capillary is a tube made up of a single layer of endothelial cells surrounded by a basement membrane. Capillaries have a lumen diameter in the range of 3–8 μm. As red cells have a diameter of about 8 μm they must deform to pass through capillaries. During passage along the 0.5–1.0 mm length of a capillary there is a volume of trapped plasma in between successive red cells. This flow pattern is known as bolus flow. Between adjacent endothelial cells there are gaps (pores or clefts) of varying size which contribute substantially to the permeability characteristics of the capillary wall. However, capillary structures are not all identical and three main types have been identified. These are designated continuous, fenestrated and discontinuous capillaries. It must be remembered that there are discrete variations in structure and function even within these three broad types of capillary. Continuous capillaries (Fig. 11.1) are the least permeable group and they are found in skin, muscle, lungs, adipose tissue and the nervous system. They are the most widely distributed type of capillary. They have tight junctions in between adjacent endothelial cells which only allow the passage of relatively small molecules (Mr<10 000). Larger molecules are thought to cross endothelial cells via the extensive invaginations (caveolae) which cover both surfaces of endothelial cells. Caveolae are linked to the formation of vesicles which can cross the cells transporting larger molecules. An intact basement membrane, which consists mainly of collagen, surrounds the endothelial tube. Pericytes partially surround the capillary and provide support. In some circumstances they are contractile and may be relaxed by nitric oxide generated in the endothelial cells (Chapter 9). Even within the classification of ‘continuous’ capillary endothelia there are considerable variations in structure and function. The least leaky capillaries are found in the brain (see p. 109). Fenestrated capillaries (Fig. 11.2A) are much more permeable to water than continuous endothelia and they are found in tissues that have a high transcapillary water flux. These include the glomerulus of the kidney, intestinal villi and the choroid plexus of the brain (the site of cerebrospinal fluid formation). There are fenestrae (round holes) in the endothelial cells which have a diameter of 50–60 nm. The fenestrae are not generally completely open holes but are covered by a very thin diaphragm which is derived from the glycocalyx. Viewed from the blood side of the capillary, these diaphragms are said to resemble a cartwheel-like structure with 14 wedge-shaped gaps. As with continuous capillaries, fenestrated capillaries have an intact basement membrane. Discontinuous capillaries (Fig. 11.2B) are found in tissues such as bone marrow, liver and spleen where there is a need for red blood cells to enter or leave the circulation. There are gaps, which may be over 100 nm in diameter, in between adjacent endothelial cells and an incomplete basement membrane which also has gaps in it. Red cells have a diameter of about 8000 nm (8 μm) but they can deform to pass through quite small holes. 1. Small lipid-soluble molecules, a group which includes oxygen, carbon dioxide and some drugs such as anaesthetic agents, can diffuse through the lipid bilayer which forms the plasma membrane of the endothelial cells. They can therefore cross almost the entire surface area of a capillary. 2. Small lipid-insoluble ions and molecules cannot easily cross cell membranes and so passage across capillary walls is confined to water-filled channels in between or through cells. There is a layer of negatively charged macromolecules (the glycocalyx) which covers the endothelial cells and lines the water filled channels. This also contributes to the permeability characteristics of the capillary wall. Small charged species (Na+, K+, Cl−, etc.) may cross capillary walls in this way but, in addition, larger hydrophilic compounds such as glucose, amino acids and many drugs also use this route. 3. Large lipid-insoluble molecules will be particularly affected by the structure of capillaries outlined above. In continuous endothelia for instance passage of these molecules will be very limited under normal conditions. In other tissues some plasma protein can leak out of the circulation and the concentration of protein in the interstitial space may be 20–70% of the plasma concentration. Many physiologically important compounds circulate bound to proteins. These include lipids, many hormones, some vitamins and micronutrients such as iron and copper. Delivery of these compounds to cells is dependent on transcapillary wall passage of their unbound form and this also applies to many drugs. A further mechanism by which proteins can cross capillary walls is via endocytotic vesicle formation. The textbook person (male, 70 kg body weight, aged 20–25 years) contains about 42 L of body water, i.e. 60% of body weight. Distribution of this water is typically about one third (14 L) in the extracellular compartment and two thirds (28 L) in the intracellular compartment (Fig. 11.3). The 14 L of extracellular fluid comprise 3 L of blood plasma and 11 L of interstitial fluid. The 2 L of red blood cells are included in the intracellular compartment. These compartments should not be regarded as ‘water-tight’ and there is a continuous flux of water between them. This process aids the distribution of nutrients around the body. Calculations based on typical values for cardiac output, and the fact that about 60% of blood volume is plasma, show that approximately 4000 L of plasma pass through the microcirculation of an adult in the course of a day (Fig. 11.4). In this period there is a net outflow of approximately 8 L of water passing across capillary walls and into the interstitial fluid. Correspondingly, 8 L of interstitial fluid drain into the lymphatic system and eventually return to the circulation by this route. Of the 8 L of fluid entering the lymph capillaries, about half is reabsorbed during passage through the lymph nodes and the other half will re-enter the circulation at the subclavian veins (see p. 132). Failure to maintain the correct distribution of body water leads to the excessive accumulation of interstitial fluid—oedema. The mechanisms behind these fluid movements are now discussed and a case history of a patient with oedema is introduced in Case 11:11. Water moves by osmosis from an area of low osmotic pressure (dilute solution) to an area of high osmotic pressure (strong solution). Osmotic pressure depends on the number of solute particles rather than their size or chemical identity. The total osmotic pressure exerted by the components of blood plasma is about 7.6 atmospheres (5800 mm Hg). In the simple model shown in Figure 11.5 this would be the pressure that would need to be exerted on top of the plasma compartment in order to just stop water moving by osmosis. Most of the osmotic pressure of plasma is generated by the small particles which have the highest concentrations in plasma, especially sodium and chloride. Capillary walls are quite freely permeable to the small particles and so the concentrations of sodium and chloride in plasma and in interstitial fluid are similar and there is no significant osmotic pressure gradient generated by the small particles. The concentrations of the small cations (Na+, K+) and the small anions (Cl−, HCO3−) each summate to approximately 150 mmol/L which means the osmolarity of plasma is approaching 300 mosmol/L. The normal range of plasma osmolarity is actually 275–290 mosmol/L. The fact that this is slightly less than 300 mosmol is partly due to electrostatic attraction between some anions and cations such as Na+ and Cl− so that a proportion are behaving as a single particle rather than two separate ones.
CAPILLARY FUNCTION AND THE LYMPHATIC SYSTEM
Structure of capillaries
Movement of substances across capillary walls
Water movement across capillary walls
Osmotic pressure of body fluids
CAPILLARY FUNCTION AND THE LYMPHATIC SYSTEM