Anatomy of the Pleura
The pleura is the serous membrane that covers the lung parenchyma, the mediastinum, the diaphragm, and the rib cage. This structure is divided into the visceral pleura and the parietal pleura. The visceral pleura covers the lung parenchyma, not only at its points of contact with the chest wall, diaphragm, and mediastinum but also in the interlobar fissures. The parietal pleura lines the inside of the thoracic cavities. In accordance with the intrathoracic surfaces that it lines, it is subdivided into the costal, mediastinal, and diaphragmatic parietal pleura. The visceral and the parietal pleura meet at the lung root. At the pulmonary hilus, the mediastinal pleura is swept laterally onto the root of the lung. Posterior to the lung root, the pleura is carried downward as a thin double fold called the pulmonary ligament.
A film of fluid (pleural fluid) is normally present between the parietal and the visceral pleura. This thin layer of fluid acts as a lubricant and allows the visceral pleura covering the lung to slide along the parietal pleura lining the thoracic cavity during respiratory movements. The space, or potential space, between the two layers of pleura is designated as the pleural space. The mediastinum completely separates the right pleural space from the left in humans. As previously mentioned, only a thin layer of fluid is normally present in this space, so it is a potential space rather than an actual one. Many diseases are associated with increased amounts of pleural fluid, however, and a large segment of this book is directed toward an understanding of these diseases.
EMBRYOLOGY OF THE PLEURA AND PLEURAL SPACE
The body cavity in the embryo, the coelomic cavity, is a U-shaped system with the thick bend cephalad. The cephalad portion becomes the pericardium and communicates bilaterally with the pleural canals, which, in turn, communicate with the peritoneal canals. With development, the coelomic cavity becomes divided into the pericardium, the pleural cavities, and the peritoneal cavity through the development of three sets of partitions: (a) the septum transversum, which serves as an early, partial diaphragm; (b) the pleuropericardial membranes, which divide the pericardial and pleural cavities; and (c) the pleuroperitoneal membranes, which unite with the septum transversum to complete the partition between each pleural cavity and the peritoneal cavity. This newly formed pleural cavity is fully lined by a mesothelial membrane, the pleura (1).
When the primordial bronchial buds first appear, they and the trachea lie in a median mass of mesenchyme, cranial and dorsal to the peritoneal cavity. This mass of mesenchymal tissue is the future mediastinum, and it separates the two pleural cavities. In humans, no communication normally exists between the two pleural cavities. As the growing primordial lung buds bulge into the right and left pleural cavities, they carry with them a covering of the lining mesothelium, which becomes the visceral pleura. As the separate lobes evolve, they retain their mesothelial covering. This covering becomes the visceral pleura in the fissures. The lining mesothelium of the pleural cavity becomes the parietal pleura (2).
HISTOLOGY OF THE PLEURA
The parietal pleura over the ribs and intercostal spaces is composed of loose, irregular connective tissue covered by a single layer of mesothelial cells. Within the pleura are blood vessels, mainly capillaries, and lymphatic lacunas. The lacunas are specialized initial
lymphatics shaped like flat cisterns and are located over the intercostal spaces, at least in sheep (3). The mean thickness of the parietal pleura in sheep is 20 to 25 µm, whereas the distance from the microvessels to the pleural space is 10 to 12 µm. Deeper to the parietal pleura is the endothoracic fascia. This continuous band of dense irregular connective tissue, composed mainly of collagen and elastin, covers the ribs and intercostal spaces and varies in thickness from 75 to 150 µm (3).
lymphatics shaped like flat cisterns and are located over the intercostal spaces, at least in sheep (3). The mean thickness of the parietal pleura in sheep is 20 to 25 µm, whereas the distance from the microvessels to the pleural space is 10 to 12 µm. Deeper to the parietal pleura is the endothoracic fascia. This continuous band of dense irregular connective tissue, composed mainly of collagen and elastin, covers the ribs and intercostal spaces and varies in thickness from 75 to 150 µm (3).
The anatomy of the visceral pleura differs markedly from that of the parietal pleura and also varies among species, primarily in its thickness. Dogs, cats, and monkeys have a thin visceral pleura, whereas humans, sheep, cows, pigs, and horses have a thick visceral pleura (4). The distinction between lungs with a thick or thin visceral pleura is important physiologically because the blood supply is dependent on the thickness of the pleura. In animals with a thick visceral pleura, the predominant source of blood is the systemic circulation; in those with a thin pleura, the predominant source of blood is the pulmonary circulation (4).
Histologically, a thick visceral pleura is composed of two layers: the mesothelium and connective tissue. Blood, lymph vessels and nerves are located in the connective tissue. Animals with a thick visceral pleura have a layer of dense connective tissue of varying thickness interposed between the mesothelium and the blood vessels (4). In sheep, the visceral pleura ranges in thickness from 25 to 83 µm (as compared with 10 to 25 µm for the parietal pleura) and the distance from the microvessels to the pleural space ranges from 18 to 56 µm (as compared with 10 to 12 µm for the parietal space) (3).
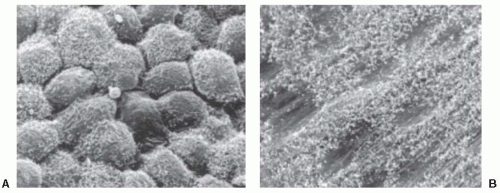 FIGURE 1.1 ▪ Scanning electron microscopic studies of the pleura. A: Bumpy pleural surface with cellular borders irregularly depressed. Note that the number of microvilli present on each cell is variable (original magnification: 1,300×). B: Flattened pleural surface with indistinct cell boundaries and sparse microvilli (original magnification: 1,250×). (From Wang NS. The regional difference of pleural mesothelial cells in rabbits. Am Rev Respir Dis. 1974;110:623-633, with permission.) |
The connective tissue layer in the visceral pleura has two important functions: (a) it contributes to the elastic recoil of the lung, which is important in expelling air from the lung, and (b) it restricts the volume to which the lung can be inflated, thereby protecting it (5). In the visceral pleura, fibers of the elastic and collagenous systems are clearly interdependent elements. Collagenous fibers are interwoven in a pleated structure that closely resembles the osiers of a wicker basket, suggesting that collagen fibers allow the lung volume to increase up to a point of maximal stretching of the system (5). The pleural contribution to the elastic recoil pressure of the lung originates from the elastic network, which returns to its resting position when inspiratory pressures are negligible (5).
Both the visceral and the parietal pleura are lined with a single layer of flat mesothelial cells. These mesothelial cells range in size from 6 to 12 µm in diameter (6). With scanning electron microscopy (7), the pleural surface is found to be either flattened or bumpy (Fig. 1.1). The bumpy areas include most of the visceral pleura and portions of the parietal pleura, including the subcostal regions and the pleural recesses. These areas appear to result from a lack of rigidity of the underlying structures (6).
Scanning electron microscopy also demonstrates that microvilli are present diffusely over the entire pleural surface (Fig. 1.1), but the distribution of the microvilli is irregular. The density of the microvilli
ranges from a few to more than 600/100 µm2, with a mean of approximately 300 (1). The microvilli are most numerous on the inferior parts of the visceral pleura and the anterior and inferior mediastinum on the parietal pleura (1). At corresponding regions in the thoracic cavity, more microvilli are present on the visceral pleura than on the parietal pleura. The microvilli are approximately 0.1 µm in diameter, and their length varies from 0.5 to 3.0 µm (1).
ranges from a few to more than 600/100 µm2, with a mean of approximately 300 (1). The microvilli are most numerous on the inferior parts of the visceral pleura and the anterior and inferior mediastinum on the parietal pleura (1). At corresponding regions in the thoracic cavity, more microvilli are present on the visceral pleura than on the parietal pleura. The microvilli are approximately 0.1 µm in diameter, and their length varies from 0.5 to 3.0 µm (1).
The exact function of these numerous microvilli is yet to be defined. At one time, it was believed that their presence increased the capacity of the visceral pleura to absorb pleural fluid. This is probably incorrect because recent observations have indicated that the visceral pleura plays a limited role in the absorption of pleural fluid. It is now thought that the most important function of the microvilli is to enmesh glycoproteins that are rich in hyaluronic acid, especially in the lower thorax, to lessen the friction between the lung and the chest wall (7). Moreover, as mentioned earlier, a thin rim of fluid normally separates the visceral and parietal pleura. Impingement of the microvilli from one pleural surface into the opposing pleural surface could possibly help maintain this thin rim of fluid (8), but this is controversial (9).
The mesothelial layer is very fragile. At thoracotomy in patients without clinical pleural disease, focal denudation of mesothelial cells is common (10). When the normal layer of mesothelial cells lining the pleura is disrupted, the defect is repaired through mitosis and migration of the mesothelial cells (11). When irritated, they retract but retain continuity with adjacent cells by projections called cellular bridges. Mesothelial cells are frequently dislodged from the pleural surfaces and are thereby free in the pleural fluid. When free in the pleural space, the cells become round or oval (11). Their cytoplasm is rich in organelles. From this state, they may be transformed into macrophages capable of phagocytosis and erythrophagocytosis (11). Such transformed cells frequently have vacuoles in their cytoplasm. Not all the macrophages in pleural fluid evolve from mesothelial cells; some definitely evolve from peripheral blood mononuclear cells, and some may evolve from alveolar macrophages (12). An immunologic role has been suggested for the macrophages derived from the mesothelial cells (12).
MESOTHELIAL CELLS
Mesothelial cells form a monolayer of specialized pavement-like cells that line the pleural surfaces. The mesothelial cells are active cells, and they are sensitive and responsive to various stimuli. The mesothelial cells that line the pleural cavity and those that line the other body cavities have no recognizable cytologic difference (13). The cytoplasm always contains a moderate to abundant amount of organelles, including mitochondria, rough and smooth endoplasmic reticulum, polyribosomes, intermediate fibrils, Golgi apparatus, and some glycogen granules, suggesting that the mesothelial cell is a metabolically active cell (14).
The mesothelium is now recognized as a dynamic cellular membrane with many important functions. These include transport and movement of fluid and particulate matter across the pleural surfaces; leukocyte migration in response to inflammatory mediators; synthesis of cytokines, growth factors, and extracellular matrix proteins; release of factors to promote both the deposition and clearance of fibrin; and antigen presentation (15). Mesothelial regeneration involves migration of cells from the wound edge and attachment and incorporation of free-floating mesothelial cells from the pleural fluid onto the denuded pleural surface (16). There is strong evidence that mesothelial cells can convert to myofibroblasts. Yang et al. (17) assessed the effects of incubating peritoneal mesothelial cells with transforming growth factor beta (TGF-β) and reported that the mesothelial cells took on the characteristic myofibroblastic phenotype. We have observed that the incubation of human mesothelial cells with TGF-β results in their morphologic transformation to cells that look like fibroblasts. It has been shown that the intrapleural administration of TGF-β results in an excellent pleurodesis (18) and the morphologic changes induced by TGF-β referred to here may be important in producing the pleurodesis.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


