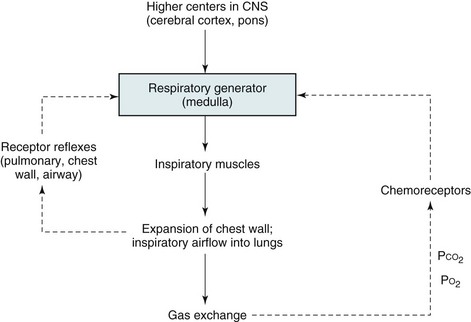
17 This chapter focuses on the anatomic and physiologic features of the controlling system and the respiratory muscles to provide background for the discussion in Chapters 18 and 19. Those chapters discuss disorders affecting respiratory control, respiratory musculature, and the chest wall. Although much of the physiology and many of the clinical problems discussed here and in the next two chapters do not directly involve the lungs, they are so closely intertwined with respiratory function and dysfunction that they are appropriately considered in a textbook of pulmonary disease. The basic organization of the respiratory control system is shown in Figure 17-1. Crucial to this system is the CNS “generator.” Signals that originate from the generator travel down the spinal cord to the various respiratory muscles. The inspiratory muscles, the most important of which is the diaphragm, respond to the signals by contracting and initiating inspiration. This process is described later in more detail under Respiratory Muscles. Maintenance of arterial blood gases is the ultimate goal of ventilatory control, and an important feedback loop adjusts respiratory center output if blood gases are not maintained at the appropriate level (see Fig. 17-1). Elevation of PCO2 (hypercapnia) and depression of PO2 (hypoxemia) both are capable of stimulating ventilation. In each case, one or more chemoreceptors detect alterations in PCO2 or PO2 and accordingly vary their input to the medullary respiratory center. The primary sensor for CO2 is located near the ventrolateral surface of the medulla and is called the central chemoreceptor. Even though it is located in the medulla, the central chemoreceptor is clearly separate from the medullary respiratory center and should not be confused with it. The central chemoreceptor does not appear to respond directly to blood PCO2 but rather to the pH of the extracellular fluid (ECF) surrounding the chemoreceptor. The pH level, in turn, is determined by the level of hydrogen (H+) and bicarbonate (HCO3−) ions as well as PCO2 (see Appendix C for further discussion). The blood-brain barrier, the permeability properties of which influence the composition of cerebrospinal fluid (CSF) and brain ECF, prevents free movement of either H+ or HCO3− from blood to brain ECF, whereas CO2 passes freely. The feedback loop for changes in PCO2 can be summarized as follows:
Anatomic and Physiologic Aspects of Neural, Muscular, and Chest Wall Interactions with the Lungs
Respiratory Control
Organization of Respiratory Control
Chemoreceptors

![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Anatomic and Physiologic Aspects of Neural, Muscular, and Chest Wall Interactions with the Lungs